 Stomata support plant survival and impact global environment Stomata support plant survival and impact global environment
A stoma (pl. stomata) is a microscopic pore on the surface (epidermis) of land plants. It is surrounded by a pair of specialized epidermal cells called guard cells, which act as a turgor-driven valve that open and close the pores in response to given environmental conditions. The presence of countless numbers of stomata is critical for plant function. Typically, the plant epidermis is tightly sealed by wax-coated, interlocking epidermal pavement cells, which protect the plant body from the dry atmosphere and UV-rays. At the same time plants must be able to breathe, or exchange carbon dioxide and oxygen, for photosynthesis and respiration. Stomata act as a gateway for efficient gas exchange and water movement from the roots through the vasculature to the atmosphere (Fig. 1). Transpiration via stomata supplies water and minerals to the entire plant system (Raven 2002)(Fig. 1). When a plant encounters adverse environmental conditions, such as drought, a plant hormone called abscisic acid triggers stomata to shut tightly in order to prevent plants from dehydration and wilting.
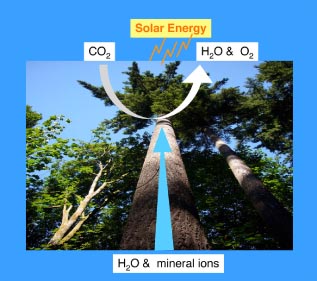
Fig. 1. Stomata as an interface of a plant and the atmosphere for global carbon and water cycle. The photograph was taken at the Keiko Torii's residence in Seattle, WA. (copy right, SJWS, 2007)
Stomatal function is important beyond the level of plant physiology and function, and its significance reaches from evolutionary history to atmospheric and environmental sciences. For instance, the acquisition of stomata is considered one of the key developmental innovations that allowed plants to conquer the terrestrial environment, an event that occurred around 400 million years ago (Edwards et al. 1998; Raven 2002). Stomata impact our present global environment as well. Hetherington and Woodward (2003) estimate that 3 x 1018 grams of carbon pass through stomata and fixed into carbohydrate every year, and in addition the total water content of the atmosphere is estimated to be recycled through stomata every half year (Hetherington and Woodward 2003). Importantly, stomata impact atmospheric environment, and in turn, atmospheric environment drives changes in stomatal patterning. It is generally accepted, both from the studies of fossil records and extant plants, that high carbon dioxide condition decreases numbers/density of stomata (Hetherington and Woodward 2003). The mechanisms that allow plants to adjust their stomatal density have just begun to unravel.
Stomatal development in Arabidopsis
Our understanding of genetic mechanisms controlling stomatal development blossomed in the 1990s, when a dramatic progress in plant biology research took place by taking advantage of a model plant Arabidopsis (Arabidopsis thaliana L. Heynh).
In Arabidopsis, stomata form anisocytic complexes, in which each stoma is typically surrounded by three surrounding cells of different sizes (Berger and Altmann 2000; Nadeau and Sack 2002). This is a relatively typical form of stomatal complexes formed in dicotyledonous plants (Esau 1977). These complexes are generated by a series of cell divisions that are stereotypical yet dynamic. During organogenesis, a protodermal cell faces a decision to undergo proliferative division to form pavement cells or undergo asymmetric division to initiate stomatal lineages (Fig. 2). A subset of protodermal cells, called meristemoid mother cells (MMC), divides asymmetrically to generate two daughter cells with distinct sizes and fates. The larger cell adopts pavement-cell identity. The smaller, triangular cell is called a meristemoid, which possesses a transient, stem-cell like property and continues a few rounds of asymmetric divisions (Fig. 2). In each asymmetric division of a meristemoid, the smaller daughter cell renews its property as a meristemoid, while amplifying the number of surrounding cells. The meristemoid then differentiates into a round, guard mother cell (GMC), which divides symmetrically once to give rise to a pair of guard cells (Nadeau and Sack 2002) (Fig. 2).
The larger daughter cells produced by the amplifying asymmetric divisions of the meristemoids are termed stomatal-lineage ground cells (SLGCs) (Fig. 2)(Shpak et al. 2005). The SLGCs resemble the pavement cells. The SLGC may divide asymmetrically to give rise to a secondary, satellite meristemoid, which differentiates into a secondary satellite stomatal complex. The SLGCs of a secondary stomatal complex may divide asymmetrically to generate a tertiary meristemoid, and so on. It is estimated that a majority of stomata (~75 %) in cotyledons and leaves is generated by satellite asymmetric divisions (Geisler et al. 2000).
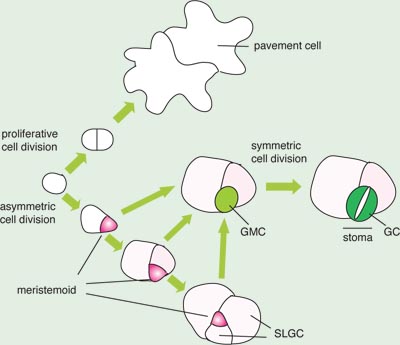 Fig. 2. Stomatal development in Arabidopsis
Fig. 2. Stomatal development in Arabidopsis
© Kyoritsu Shuppan, 2006
References
Berger, D. and Altmann, T. (2000) Genes & Dev. 14: 1119-1131
Edwards, D., H. Kerp, et al. (1998) J Exp Bot 49: 255-278
Esau, K. (1977) Anatomy of seed plants. John Wiley & Sons, New York: pp88-99
Geisler, M., J. Nadeau, and F.D. Sack (2000) Plant Cell 12: 2075-2086
Hetherington, A. M. and Woodward, F.I. (2003) Nature 424: 901-908
Nadeau, J. A. and Sack, F.D. (2002) Stomatal Development in Arabidopsis, In Arabidopsis Book, ASPB.
Raven, J. (2002) New Phytol 153: 371-386
Shpak, E.D., McAbee, J.M., Pillitteri, L.J., and Torii, K.U. (2005) Science 309: 290-293
copy right, Keiko U. Torii, 2006
|