 ERECTA, a putative receptor kinase regulating plant size ERECTA, a putative receptor kinase regulating plant size
Genetic and molecular evidence suggests
that the Arabidopsis ERECTA mediates cell-cell communication that is
required for coordinated cell proliferation and organ growth.
Loss-of-function erecta mutations confer reduced plant size and compact
stature. The effects of erecta mutations are quantitative. In other
words, more severe the mutation is, smaller the plant size will be
(Fig.1). The erecta phenotype, including compact inflorescence with
short lateral organs and internodes, is largely due to reduced cell
numbers in the cortex cell files (Fig.2). The shortened
erecta pedicels (floral stems) are associated with an increase in 4C
cells, suggesting possible aberration in cell cycle progression (Fig.2).
We have cloned the ERECTA gene some years ago, and we found that
it encodes a leucine-rich receptor-like serine/threonine kinase
(LRR-RLK), a prevalent subfamily of signaling receptors in plants (Torii
et al., 1996). As a collaboration with Prof. John Walker’s lab (Univ.
Missouri), we demonstrated that ERECTA has a functional serine/threonine
kinase activity (Lease et al., 2001).

Fig. 1.
The degree of defects in ERECTA directly transltaes to the plant size.(from right) wild-type ERECTA, weak er-116, intermediate er-103, and KO er-105
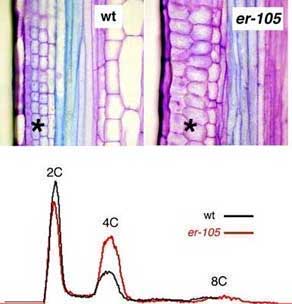
Fig. 2.
ERECTA is required for coordinated proliferation in the pedicel cortex cell files (*). er-105 has a short pedicel with reduced cell numbers, that are highly disorganized. (bottom)The erecta mutation confers an increase in 4C nuclear content, suggesting potential defects in cell cycle.
Redundancy in the ERECTA pathway revealed by a dominant-negative approach

Fig. 3.
Exression of a dominant-negative DKinase fragments driven by the native ERECTA promoter severely enhances growth defects of erecta KO plants.
How does ERECTA act as a
receptor? How does ERECTA-signaling pathway integrate cell-cell signal
to cell proliferation in the context of organ growth? As a first step to
address the questions, we decided to analyze its mode of action. For
this purpose, we generated transgenic Arabidopsis plants that express
distinct domains of ERECTA. We found that truncated ERECTA that lacks
the cytoplasmic kinase domain (Kinase) acts as a dominant-negative receptor. The transgenic wild-type plants expressing DKinase
fragments are phenotypically identical to erecta mutant plants. More
strikingly, the expression of the dominant-negative ERECTA fragment
severely enhanced the growth defects of the erecta knockout allele (Fig.
3). That having truncated ERECTA protein is worse than having none at
all for Arabidopsis organ and internodal growth suggests redundancy in
the ERECTA- signaling pathways (see Fig. 4 for our model). The DKinase
fragments may take up and deplete ligands for other receptors.
Alternatively, the fragments may directly interact with shared receptor
partners of ERECTA, and thus shut down whole pathways. Consistent with
this hypothesis, we found that a functional LRR domain is required for
dominant-negative interference (Shpak et al.,2003).
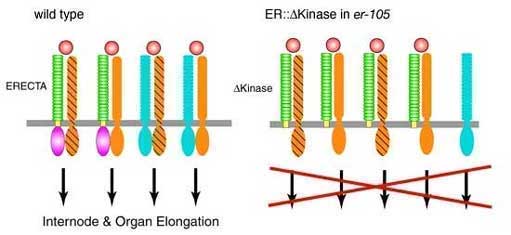
Fig. 4.
Model for the ERECTA signaling pathway and DKinase action.
Synergistic interaction of three, paralogous ERECTA-family RLKs
Our dominant-negative study of
ERECTA revealed the presence of redundancy in the ERECTA-mediated signal
transduction pathway. We went further ahead and identified redundant
receptors, ERL1 and ERL2 (cyan molecules in Fig. 4. Shpak et al. 2004).
They are functional paralogs of ERECTA evolved by recent gene
duplication (Fig. 5). ERECTA, ERL1, and ERL2 are ‘functionally
equivalent’ receptors, because expression of ERL1 and ERL2 driven by the ERECTA promoter rescues growth defects of erecta
KO plants (Fig. 6). In other words, ERL1 and ERL2 are capable of
perceiving and transducing the same signal as ERECTA. Overlapping but
unique expression patterns of ERL1 and ERL2 make these two receptors
distinct from (but partially redundant with) ERECTA.
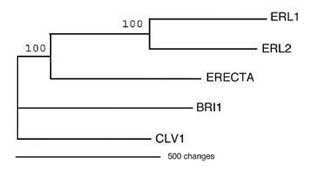
Fig. 5. ERL1 and ERL2 are immediate paralogs of ERECTA arisen by recent gene duplication. Shown in a most parsimonous tree.

Fig. 6. ERECTA, ERL1 and ERL2 are functinally equivalent. ERL1 and ERL2 genes rescue the erecta phenotype when expressed under ERECTA promoter.
We subsequently identified KO mutants of ERL1 and ERL2 to understand their roles in plant growth and development. erl1 and erl2
are redundant, that is, they conferred no detectable phenotype by their
own, but they enhanced erecta defects in a unique manner. Loss of the
entire ERECTA family genes led to striking dwarfism, reduced
lateral organ size, and abnormal flower development, including defects
in petal polar expansion, carpel elongation, and anther and ovule
differentiation (Fig 7). These defects are due to severely reduced cell
proliferation. Our findings place ERECTA-family RLKs as redundant
receptors that link cell proliferation to organ growth and patterning.

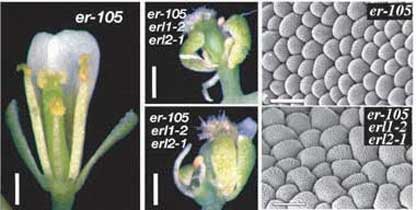
Fig. 7. ERECTA, ERL1 and ERL2 interact synergistically to promote plant growth and development. The erecta erl1 erl2
triple mutant plants are severely dwarfed and develop abnormal flowers
due to defective cell proliferation. (Bottom right) Two scanning
electron micrographs showing the petal epidermal cells. As you can see,
the triple mutant petals are small and needle like, but their epidermal
cells are large and disorganized, indicating severe defects in
coordinated cell proliferation.
© Keiko U. Torii, 2004
|