
Kollman Lab
Research
Kollman Lab
Research
Assembly and organization of magnetosomes
A phylogenetically diverse group of magnetotactic bacteria (MB) are able to orient themselves in magnetic fields, which is thought to simplify their search for optimal growth conditions. MB orient themselves by creating intracellular magnetic crystals. This tightly controlled process of biomineralization occurs in small (~50 nm) organelles called magnetosomes formed from invaginations of the inner membrane that remain open to the periplasm. Because of their small size and the tractable number of genes involved in their formation, magnetosomes have become an attractive model system for understanding basic principles of organelle biogenesis. The ability to control biomineralization of magnetic nanocrystals also makes magnetosomes potentially useful in biotechnological and medical applications.
Recent genetic, proteomic, structural and cell biology studies have defined many of the magnetosome components, and indicated functions for a subset of these. The magnetosome proteome consist of both integral membrane and soluble proteins, many of which are encoded by gene clusters within a large genomic region named the magnetosome island. In particular, the mamAB gene cluster is essential for magnetosome formation and organization and magnetite biomineralization. A systematic genetic dissection of the mamAB genes revealed a stepwise assembly process of the magnetosome. Magnetosomes are aligned by the actin-like filament MamK into a regularly spaced chain along the length of the cell. Only after the organelles have been formed and aligned does the process of biomineralization begin.
Our focus is on the formation of the membrane-bound magnetosome structure and its organization within the cell. While there is now a wealth of information about the proteins involved and the overall morphology of the organelle system, little is known about the molecular mechanisms of magnetosome assembly. How is the inner membrane initially deformed? What maintains the opening between the magnetosome compartment and the periplasm? With an opening to the periplasm, how is a distinct environment maintained in the magnetosome? How are magnetosome-specific proteins targeted to the organelle? How does attachment to MamK filaments mediate alignment, and how is the spacing between aligned magnetosomes maintained? How is magnetite crystallization initiated and controlled, and how is the crystal attached to the membrane?
To begin to answer these questions, we are generating a model of magnetosome structure and alignment that will serve as a framework for both interpreting and driving new cell biological and biochemical experiments. Our overall approach is to determine structures of isolated magnetosomes by cryo-electron tomography, and combine this with higher resolution structures of magnetosome components determined by X-ray crystallography and cryo-EM to generate an integrated model of the magnetosome.
magnetosome chain in
M. magneticum AMB-1
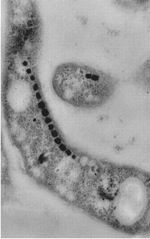
empty magnetosomes from whole cryo-ET

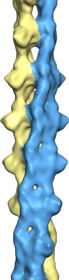
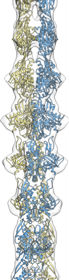
cryo-EM reconstruction
of MamK filaments
1959 NE Pacific Street Box 357350
Seattle, WA 98195