A further example is the analysis of hourly increments of conifer shoot growth using a model of environmental variables by Rie Komuro.
In both examples the model was improved and choices of assessment criteria were developed as model assessment took place.
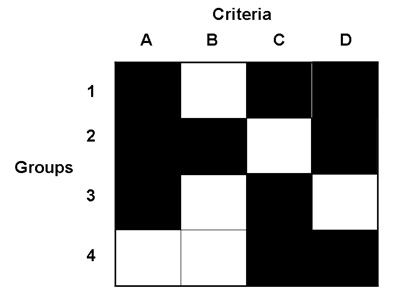
Illustration of a set of assessment vectors where four criteria, A, B, C and D, are not satisfied by any one group. Satisfied criteria are represented by black squares, unsatisfied criteria by white squares. A group contains at least one parameterization of the model that achieves all of the criteria marked as black squares. Individuals in groups 1 and 2 both achieve three criteria and are non-dominated but those in group 3 and group 4 are both dominated by group 1. Some parameterizations of the model may not achieve a unique group of criteria and are considered as "dominated" by those that do and are discarded from further consideration. The evolutionary computation algorithm continually attempts to produce groups that achieve more combinations of assessment criteria.