|
1. Cone photoreceptors
provide color vision as well as high acuity vision
Seeing things in color is something we take for granted, unless we know
someone who is colorblind and we become aware of the difficulties this
presents. Color vision is not only a pleasurable experience, it also
helps us locate and identify objects in the visual scene that would be
hard to pick out in shades of gray. The ability to discern different
wavelengths, or colors, of light gives us more information for detecting
and identifying objects than would be provided solely by black and white
vision, as demonstrated in Figure 1.

Figure 1. The same geometric drawing in color and in shades of gray.
(Image courtesy of Webvision)
Have you noticed that as daylight fades or as room lights dim, colors
become hard to identify? Color vision is mediated by specialized nerve
cells in the retina called cones, which function only in
bright light. When light becomes dim, rods take over, and
these provide neither color vision nor high acuity (ability to detect fine
detail, such as that needed for reading). Cones and rods are types of
photoreceptors.
2. Several different
pigments are needed for color vision
How do cone photoreceptors begin the process by which we perceive colors?
As discussed in an earlier unit (see the Background
section for Our Sense
of Sight, Part 1), both rods and cones contain a visual
pigment. This pigment molecule, which is embedded in the cell
membrane of the photoreceptor, is a large protein called an
opsin, and it is coupled with a small molecule called a
chromophore (a form of Vitamin A) that absorbs photons of
light. When the chromophore absorbs light, it changes shape, and this in
turn activates the large opsin molecule. In domino-like fashion, this
leads to a cascade of molecular events that culminates in an electrical
change in the cell membrane. The electrical signal is transmitted to
retinal ganglion cells, whose axons take the information about light, now
encoded as electrical signals, to the brain.
If all photoreceptors had the same pigment, however, we could not
distinguish different wavelengths (i.e., colors) of light. To understand
this, consider light of a given wavelength, say somewhere in the green
region of the spectrum, falling onto a photoreceptor cell. The cell gives
a certain response, perhaps 20 arbitrary units. If a blue light of the
same intensity strikes, the cell may give a response of 10 units, because
the blueness gives it a different quality from the green light. But if
the intensity of the green illumination is reduced to the point where the
cell gives a response of 10 units, the cell will code this input the same
as a brighter blue light: the color information is unclear. Because
photoreceptors, like all neurons, can only signal a change by increasing
or decreasing their electrical output, the signals are ambiguous as to
whether the change in response (here, 10 units vs. 20) is due to a
difference in wavelength or in intensity. So with only one visual
pigment, we would see the green light that gave a 20 unit response as
brighter than the blue, but we would not see them as different colors.
Researchers tell us that, in principle, three pigments, or three types of
photoreceptor cells with different combinations of three pigments, could
produce vision capable of detecting all colors of the visible spectrum
(400 to 700 nanometers). Scientists have indeed found that the human
retina (and a few others) has three types of cones, each containing a
different visual pigment. These are the red, green, and blue cones, each
containing a pigment similar to rhodopsin, the pigment found in rods.
Each cone pigment is an opsin, but the opsins have different amino acid
sequences that restrict the accompanying chromophore (in a yet unknown
manner) so that it preferentially absorbs only one part of the visible
spectrum. Thus, red cones absorb longer wavelengths, green cones medium
wavelengths, and blue cones shorter wavelengths.
Each type of cone cell responds to a range of
wavelengths rather than just
one, and their response curves overlap (Figure 2). Because of this
overlap, each wavelength of light gives a unique pattern of response in
each of the three cone populations, and the pattern of signals is the code
for the wavelength, or color. That is, the color is encoded in the
pattern of activity of a number of cells rather than in the isolated
activity of one cell. This mechanism is similar to that for olfactory
perception: individual olfactory receptors give different responses to the
same odor, and the odor is coded by the pattern of receptor activation.
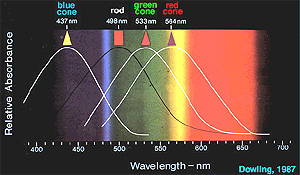 |
Figure 2. The wavelength sensitivities (response curves) of the
different photoreceptor types in the vertebrate retina. (Image courtesy
of Webvision)
|
3. Some animals have
color vision that differs from that of humans
Many animals do not have color vision: the ability to discern different
wavelengths evolved in animals that became diurnal, or active in the
daytime. Among these, birds are especially color sensitive. Some have
four or more cone pigments, allowing them to distinguish more accurately
among colors than we do (probably seeing more shades), and to see into the
ultraviolet range, which we cannot.
Most primates other than humans have just two visual pigments, one for
short (blue) wavelengths, and one that varies in different animals but
detects light somewhere in the red to green part of the spectrum. Familiar
animals such as dogs and cats also have two types of cones. Old World
monkeys have three cone types, as humans do.
Some invertebrates have color vision: like birds, some insects can see
ultraviolet light. Insects also have different screening materials in the
corneal covering of their eye facets; this allows for tuning different
eye units (called ommatidia) to different wavelengths.
4. Color signals travel from the retina to the
thalamus and on to the visual cortex
Once the cones have generated a pattern of responses to a color stimulus,
the local circuits of neurons within the retina (Figure 3) process the
information before sending it to the final retinal station, the ganglion
cells. These circuits abstract and enhance cone signals, beginning the
neural processing that allows us to recognize differences in color despite
wide variations in light levels. (Keep in mind that color information is
only one aspect of vision that cones are responding to. They are also
responsible for all other aspects of vision in bright light, including
form and motion. This information is processed in separate neural
circuits in the retina and brain.)
| Figure
3. Schemata of the cells of the retina. This drawing shows some of the
important cells in retinal circuits; the actual connections are much more
complex. (Image courtesy of Webvision) |
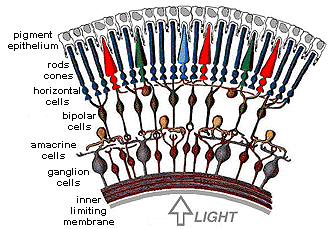 |
Like other visual information, signals coding color travel from the
retina, via axons of the ganglion cells, to the lateral geniculate nucleus
of the thalamus. Axons from color-processing cells in the geniculate are
connected to color-detecting cells in the visual cortex, in the occipital
lobe at the back of the brain (Figure 4).
Within the occipital lobe, cells in the primary visual cortex, often
called "area 17" by researchers, are the first to receive messages from
the geniculate. Signals conveying color information then go on to several
nearby visual areas for further processing, mainly to an area termed "V4."
For perception and recognition, signals are then sent to so-called "higher
centers," where they interact with stored memories and input from other
sensory and motor centers. One "set" of information is spatial and goes
mainly to the parietal lobe, while a second "set" concerned with object
recognition and color perception goes to the lower part of the temporal
lobe. However, scientists have also found that even when these parietal
and temporal lobe areas are damaged, people can perform tasks involving
spatial relationships, object recognition, and color information, so the
brain must also process this visual data in additional places.
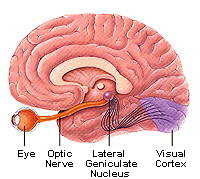 | Figure 4. The
visual pathway. Axons from ganglion cells in the retina travel in the
optic nerve to the lateral geniculate nucleus of the thalamus. Here they
synapse with other neurons, whose axons go to neurons in the visual cortex
in the occipital lobe of the brain. (Figure courtesy of the Society for
Neuroscience, copyright by Lydia Kibiuk, 1994.) |
5. The "opponent-color"
theory helps explain some features of color vision
Scientists are able to explain some curious features of color vision, even
though all of the mechanisms are not yet fully understood. These features
include successive color contrast, simultaneous color contrast, and color
constancy. The "opponent-color theory" helps explain these phenomena. The
following summaries are quite sophisticated and should be simplified for
younger middle school students.
Researchers studying color perception noted that certain colors could not
be seen in combination; for example, we do not see colors called
"red-green" or "blue-yellow." Therefore, these colors, along with black
and white, were described as opponents or antagonists. Scientists also
observed that a certain combination of red (long) and green (medium)
wavelengths produced the color yellow. They could verify that this
combination would be equivalent to a third wavelength, but could not
explain why this combination looked yellow instead of "reddish-green."
The opponent-color theory formulated in the 1960s, accounts for some of
these observations. Researchers believe that, starting in the retina,
information about color and about intensity of light is sorted into three
"channels." The channels consist of axon pathways from retinal ganglion
cells, which receive all cone information, to the brain. Two of these
channels carry color or wavelength information, and one carries intensity
information-the degree of blackness or whiteness.
One of the two color channels responds to red or to green light: here,
certain ganglion cells will fire signals if stimulated by red light
(messages sent by red cones) and will decrease firing if they get signals
from green cones; other ganglion cells do the opposite. In the other
channel, blue-yellow ganglion cells work the same way. The "intensity
channel" also works in a similar manner: here, the sum of red, green, and
blue cone input results in different ganglion cells detecting black,
white, or shades of gray. (Remember that if all the colors of the visible
spectrum are summed, they appear white.)
As shown in Figure 5, each ganglion cell receives input from a defined set
of cones, and when a ganglion cell gets the necessary signal combination
to activate it, it sends a message that indicates a particular color.
Thus, a "blue" ganglion cell, one that only signals "blue" to the brain,
fires a message when it receives input from blue cones but no input from
green or red cones. A "yellow" ganglion cell sends its "yellow" message
to the brain when activated by a certain pattern from green and red cones,
but no messages from blue cones. Remember that while yellow is signaled
by activation of both red and green cones, the message "yellow" travels in
the same pathway, or nerve fiber bundle, as the message "blue," and these
two (yellow and blue) cannot travel at the same time, so we do not see
"blue-yellow." The messages "red" and "green" are sent by different
ganglion cells from those that send "yellow," even though all these are
receiving information from (different) red and green cones. Because red
and yellow are transmitted in separate channels, we can simultaneously see
red and yellow (orange), and for the same reason, we can see red and blue
(magenta), blue and green (aqua) and yellow and green (lime).
 | Figure
5. Simplified diagram (after McIlwain, 1996) of how colors might be
perceived based on the opponent-colors theory. See text for explanation.
This diagram ignores the internal retinal circuits. |
For younger students:
Try comparing the opponent color channels to tubes that can only carry one
kind of marble at a time. For example, one tube carries both red
marbles (red light) and green marbles (green light), but only one type can
travel through the tube at a time. When red marbles come out the end of
the tube, they hit a switch in the brain and turn it on, signaling red,
while the green marbles signal green. Because only red or green can
travel at one time, we cannot see colors that are combinations of red and
green ("red-green"). Blue and yellow marbles would have their own
separate tube, so we can see blue and green, or red and yellow, at the
same time: both tubes can work simultaneously. This explanation leaves
out the fact that yellow is a combination of red and green wavelengths-the
yellow marbles are a simplification.
Successive color contrast: the color afterimage
If you look at a bright red circle for a time and then at a gray or white
background, you see a green circle or afterimage (and
similarly, you see a red afterimage for an initial green object.) The
opponent-color theory helps explain this phenomenon. Prolonged viewing of
the red stimulus causes adaptation in the red-selective cones: like other
primary sensory cells, they stop firing if bombarded for too long with a
stimulus. When the retina is subsequently exposed to a white light
(reflected from the white or gray background) the red cones are unable to
fire for a short time, and the ganglion cells connected to these cones
decrease their firing. The green cones are not
fatigued, so the ganglion cells receiving green light send messages
through the red-green channel, and the brain receives only a green signal.
For the red-green channel to signal "white," it must receive signals from
both red and green, which then "cancel" each other. Other pairs of colors
will also give color afterimages: blue and yellow, and black and white.
Further, different shades of initial colors will give different shades of
their afterimage colors.




Figure 6. Afterimages: Stare at the yellow + in the middle of the figure
on the left for 15-30 seconds. Then move your gaze to the white square.
Did the colors reverse themselves? Try the same with the red and green
figure. These are examples of "afterimages." See text for
explanation.
Simultaneous color contrast
Another visual phenomenon is observed when a gray object seen against a
background of green has a red tinge, and vice versa. The mechanism also
begins in the retina, where messages are funneled into their appropriate
channels, but the explanation is more complicated. The information is
transmitted to special cells in the visual cortex called "double opponent"
cells. These cells respond best to opponent colors in a certain spatial
arrangement, such as a red spot in a green background, or the reverse.
Researchers think this response characteristic of the visual cortical
cells helps produce the green tinge seen with a gray object in a red
background: the double opponent cells "fill in" the green circle because
that is what they are "programmed" to see. This phenomenon is not yet
fully understood.
Color constancy
Researchers also believe that double opponent cell activity is responsible
for the fact that an object seems to remain the same color in spite of
changing light conditions, say, from bright daylight to dusk, or from
sunlight to artificial light. This has to do with the fact that to a
great extent, we judge color by comparing the object and its background:
the entire retinal image is taken into account by our brains. However, we
have all had the experience of buying an article of clothing that looks
one color in the artificial lighting of a store, but appears to
be a different shade in sunlight. In this case there is no constant
background for comparison.
In a further apparent exception to the color constancy rule, a set of
objects reflecting identical wavelengths from their surfaces (for example,
red light-they should all look red) can appear to be completely different
colors if set against different backgrounds, or when using different
ambient lighting conditions.
6. Defects in color vision are usually
genetic
Although we speak of "color blindness," it is extremely rare for a person
to see no colors at all; rather, these people usually either cannot
detect one color, or they perceive it differently from the normal
population. Researchers have determined that these defects are generally
genetic, and involve the photoreceptor pigment genes.
The genes for the red and green pigments are both on the X chromosome,
quite close to one another, and because men have only one X chromosome,
they are more likely to be missing one gene or to have a recombined and
thus abnormal gene; approximately 2% of men are red-green colorblind.
Mutations in blue cone pigment genes are unusual: this gene is on
chromosome 7, so everyone has two copies and both would have to be
defective in order for a problem to occur. Approximately eight percent of
males, and fewer than one percent of females, have some difficulty with
color vision.
 |
Figure 7. A test for red-green colorblindness. People with normal
color vision should see an 8 on the left and a 5 on the right. People with
red-green color blindness may see 3 on the left 2 on the right. Note that
tests displayed on color monitors may not produce accurate results. If
you think that you are having trouble seeing colors correctly, have your
color vision checked by a doctor. These figures were originally devised
by Dr. Shinobu Ishihara (Ishihara, S., 1954. Tests for colour-blindness.
Tokyo: Kanehara Shuppan.) |
Acquired defects in color vision can be caused by lesions (such as
hemorrhages from stroke or accident, or tumors) in area V4 of the visual
cortex. These must usually occur on both sides of the brain to affect
color vision.
|

 [Back to Top]
[Back to Top]![[email]](./gif/menue.gif)

