
Mass extinctions played a dual role in influencing evolution: culling species and their evolutionary traits and generating open ecospace and new opportunities for survivors (Gould 1985, Raup 1994). The outcomes have dramatically restructured the biosphere and redirected the course of evolution. Our ongoing work on the Cretaceous-Paleogene (K-Pg) boundary examines the patterns and processes of mass extinctions and the biotic recoveries that follow.

The K-Pg extinction
Substantial evidence supports an extra-terrestrial impact at the K-Pg boundary, but the causes, timing, and extent of the associated mass extinction remain unclear. The K-Pg boundary section in northeastern Montana is a standard for examining patterns in the terrestrial realm, but few studies have examined the biotic changes at ecologically relevant spatial and temporal scales leading up to, across, and following the K-Pg boundary.
We have compiled a large fossil database of latest Cretaceous and early Paleocene vertebrate faunas from the Hell Creek Formation and Tullock Member of the Fort Union Formation of northeastern Montana. It includes vertebrate microfossils collected by our field crews over the past 16 years, and those amassed during the 30+ years of collecting prior to our involvement (Archibald 1982, Clemens 2002, Wilson 2004, 2005, Clemens and Wilson 2009).
Fossil localities have been integrated into a temporal framework based on stratigraphic position. For example, changes in mammal, turtle, and amphibian paleo-communities leading up to the K-Pg boundary have been compared to published floral and paleoclimatic data from the same region.
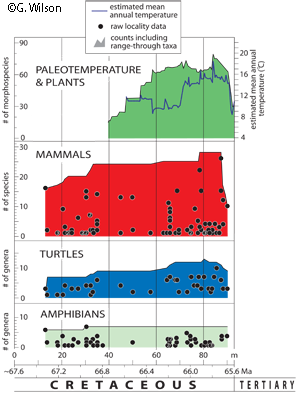
Results show no evidence of long-term changes in richness leading up to the K-Pg boundary, although changes in relative abundances, taxonomic composition, and body sizes could reflect destabilization in the 300 to 500 thousand years leading up to the K-Pg (Wilson 2014, Wilson et al. 2014). These local biotic changes are coincident with declining temperatures locally (Tobin et al. 2014), broadly coincident with a major pulse in Deccan volcanism in India, and declines in marine biotic diversity. More dramatic changes in mammal and turtle paleocommunities occur at or near the K-Pg boundary
The results are consistent with a multiple-cause hypothesis for the mass extinction; rather than a single cause in the form of the bolide impact. Future work will improve the temporal resolution of the chrono-stratigraphic framework, further develop the local climate signal, improve the density of fossil sampling, incorporate other fossil taxa (plants, molluscs), and draw comparisons with other study areas.
Paleocene ecological expansion of placentals
It has long been held that following the K-Pg extinction of non-avian dinosaurs, placental mammals rapidly invaded a wide array of ecological niches via in situ evolution and immigration. The predicted pattern of increasing species richness of placentals has been broadly documented (e.g., Stucky 1990, Alroy 1999), but the pattern of the morphological diversification expected from the ecological diversification has not been directly quantified.
I am tracking morphological patterns of mammal paleocommunities across the K-T boundary to infer the pattern and timing of ecological expansion in placental mammals after the K-T boundary. Specifically, I have applied geometric morphometrics to therian molars and multituberculate fourth premolars to quantify changes in the dental morphological disparity and morphospace occupation of mammal paleocommunities across the K-T boundary in the Hell Creek study system (Wilson 2003, 2004). I have also estimated changes in body size in these paleocommunities.
The results suggest that dental morphological diversification of placental mammals lagged several hundred thousand years behind taxonomic diversification, which immediately followed the K-Pg boundary (Wilson 2004, in prep.). This research may have implications for understanding evolutionary and ecological processes, like adaptive radiations and recovery from mass extinctions. Future work will track these patterns in younger time horizons and compare them to patterns from other areas, like the Denver Basin. My colleagues and I are also developing new methods for quantifying tooth shape and its relationship to diet (Wilson et al. 2006, Evans et al. 2007), which will further refine these patterns across the K-T boundary.
Collaborators: William A. Clemens, Harley J. Garbani, Michael Poltenovage, Grace Carter, David DeMar, Jr.
Funding: UW Royalty Research Fund
Publications: Wilson, G.P. 2005. Mammalian faunal dynamics during the last 1.8 million years of the Cretaceous in Garfield County, Montana. Journal of Mammalian Evolution 12(1/2):53-76.
Clemens, W.A. and G.P. Wilson. 2009. Early Torrejonian mammalian local faunas from northeastern Montana, U.S.A. Papers on Geology, Vertebrate Paleontology and Biostratigraphy in Honor of Michael O. Woodburne. Albright, L.B. III (ed). Museum of Northern Arizona Bulletin 65:111-158.
Archibald, J.D., W.A. Clemens, K. Padian, T. Rowe, N. MacLeod, P.M. Barrett, A. Gale, P. Holroyd, H. Sues, N.C. Arens, J.R. Horner, G.P. Wilson, M.B. Goodwin, C.A. Brochu, D.L. Lofgren, S.H. Hurlbert, J.H. Hartman, D.A. Eberth, P.B. Wignall, P.J. Currie, A. Weil, G.V.R. Prasad, L. Dingus, V. Courtillot, A. Milner, A. Milner, S. Bajpai, D.J. Ward, A. Sahni. Cretaceous Extinctions: Multiple Causes. Comment. Science 328:973.