Research Overview |
|
I am broadly interested in predator ecology, and especially the role played by these species in communities as agents of prey mortality and behavioral modification (i.e., in predator effects). I also maintain an active interest in the ecology of snowshoe hares and other leporids, with special reference to their interactions with predators. My research program pursues these interests using a combination of field and laboratory work in marine and terrestrial systems (see list of publications). Some specific research topics on which my program is currently focused are listed below. |
|
Contingency in Predator Risk Effects |
|
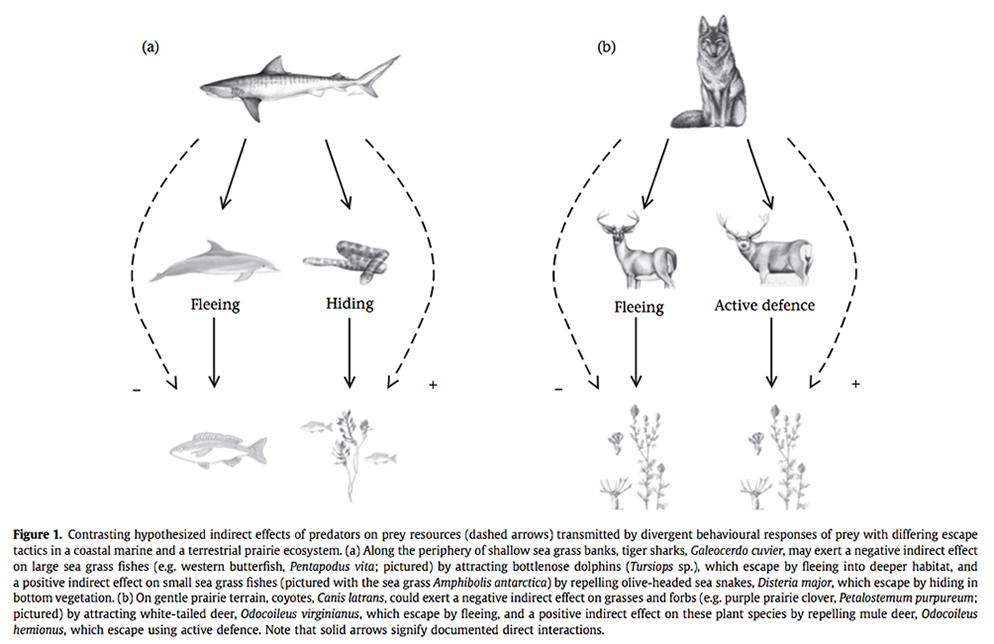
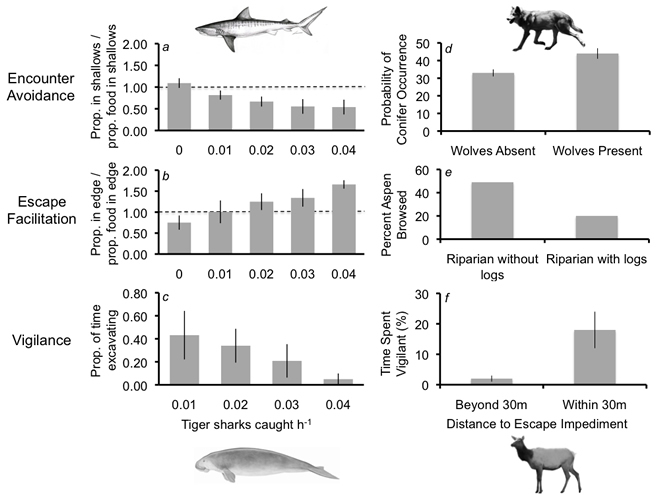
Prey often modify their behavior in response to the threat of predation. Such anti-predator behavior can alter patterns of resource exploitation by prey and ultimately trigger changes to prey populations and community properties (i.e., risk effects). The implications of risk effects for particular communities, however, have proven difficult to anticipate in large part because anti-predator behavior is not generalized. Rather, prey responses to predation risk appear to be highly contingent, or context dependent. One of the main goals of my research program is to explore the sources of this contingency, asking whether and to what extent risk effects in particular cases hinge on predator hunting modes, prey escape tactics, and the setting of the predator-prey interaction (i.e., landscape context). My graduate students and I will address these questions by focusing on interactions between tiger sharks (Galeocerdo cuvier) and large marine consumers (e.g., dugongs, Dugong dugon) at my long-term field research site in Shark Bay, Western Australia and between a variety of carnivores and their prey in the Pacific Northwest of the United States (e.g., between gray wolves, Canis lupus, and cervids). Our goal is to help develop a general framework that uses the drivers of behavioral contingency to better understand and presage risk effects and their community consequences (see figure from Wirsing et al. 2010 below). The development of such a framework is important for conservation, for it will enable more reliable prediction of the ecological impacts of losing predators from, and repatriating predators to, ecosystems. |
|
|
 |

*Photographs by Michael Heithaus