Enzymology's New Frontiers
Investigators are learning about structures and rethinking
old theories | By Mark
Greener
"There's no big mystery to being an enzymologist. All you
have to have is a razor blade and a liver."
--Gordon Tomkins to Julius Axelrod, circa
1950
Today's researchers rarely resort to razor blades to
understand how enzymes function. Investigators can capture the
kinetics of a single enzyme molecule or use site-directed
mutagenesis and crystallography to appreciate the subtleties
of enzymatic action.
Such technological advances help enzymologists understand
the fundamental biological processes that have an impact on
life sciences as diverse as systems biology, neurology, and
ecology. Modern enzymology also has numerous commercial
applications, which include a plethora of new pharmaceutical
targets.
The field also has an abundance of unsolved, pressing
problems: many of the current frontiers in enzymology require
multidisciplinary approaches. In particular, the field needs
to evolve away from examining enzymes in isolation.
Increasingly, enzym-ology aims to understand entire
biochemical pathways and the way in which genotypic and
proteomic variations influence outcomes.
"There is no doubt in many people's minds that there is, or
will be, a renaissance in enzymology as a discipline," says
Hong Qian, associate professor at the Department of Applied
Mathematics, University of Washington, Seattle. "But it will
study many enzymes in a complex biochemical reaction network,
not just one enzyme at a time." As a result, many
enzymologists are moving away from relatively simplistic
deterministic kinetics and toward sophisticated, probabilistic
mathematics and nonequilibrium thermodynamics.
A recent study by research fellow Kirsten Donald,
Department of Zoology, University of Otago in New Zealand, and
colleagues1 underscores the power of merging
traditional enzymology and new technology in the study of
biological systems. The investigators combined quantified gene
expression using fluorogenic real-time PCR and classic
enzymatic assays to understand the role and regulation of
proteolysis in the early development of the Pacific oyster
Crassostrea gigas. "Measuring both expression and
activity over time allowed us to understand how
protein-turnover enzymes were regulated," she remarks. Some
enzymes, such as leucine aminopeptidase, are regulated at the
transcriptional level. Others, including cathepsin B, undergo
posttranscriptional modification.
UNDERSTANDING STRUCTURES Full structural analysis of
the enzymes in the pathway is another landmark of the systems
biology frontier and a multidisciplinary endeavor.2
Indeed, recent studies using site-directed mutagenesis and
crystallography have shown researchers the finer details of
enzymatic action. Thomas Dandekar, from the European Molecular
Biology Laboratory in Heidelberg and the Department of
Bioinformatics at the University of Würzburg, Germany,
suggests that such findings could help inform "nearly all
biological disciplines" including microbiology, immunology,
neurobiology, and ecology.
Crossing this frontier means moving beyond genomics;
sequence homology may not offer an appropriate surrogate for
structure. Even proteins with more than 80% homology can show
different structural features in the active domain that may
influence specificity, selectivity, and
reactivity.3 Against this background,
crystallography becomes increasingly important. "We have
learned so much about how enzymes work by the analysis of
their structures," says professor Gail Johnson, Department of
Psychiatry at the University of Alabama, Birmingham.
By 2000, researchers had structurally characterized some
640 enzymes, including, for example, the enzymes involved in
the glycolytic pathway.3 Since then, the number of
characterized structures has tripled. The BRENDA database (http://www.brenda.uni-koeln.de/),
maintained by the University of Cologne, Germany, collates
data on the 35,000 or so enzymes classified according to the
Enzyme Commission nomenclature. The database now contains some
1,900 crystallography results as well as the tertiary
structures of 11,000 enzymes.
However, the number of structural characterizations should
increase markedly over the next few years. Traditionally,
determining a protein's 3-D structure using X-ray
crystallography took months or even years. But technological
advances that integrate robotic systems and new software for
data collection and processing now produce fully automated,
high-throughput crystallography systems, which can determine
the structures of 18 crystals of aldolase and 54 crystals of
protein tyrosine phosphatase 1B in 42 and 80 hours,
respectively.2
This growing structural database holds some surprises. For
instance, researchers recognized years ago that tissue
transglutaminase bound and hydrolyzed guanine triphosphate
(GTP). But no domain showed homology with any other known G
protein. Indeed, mutational studies suggested that the GTP
binding and hydrolysis domain resided in the N-terminus. In
2002, crystallography of tissue transglutaminase revealed a
unique GTP binding and hydrolysis domain in the
C-terminal.4 As Johnson remarks, "You never know
until you see the structure!"
Site-directed mutagenesis augments crystallography's
revelations. For example, it allows researchers to substitute
cysteine with serine. This retains the residue's size and
chemical character and, therefore, does not disrupt protein
folding. Few such substitutions lead to a total loss of
activity. "This suggests that active sites of enzymes are
optimized for the catalytic step and promote catalysis even
when the key residue is knocked out," comments Wilbur
Campbell, emeritus professor of biochemistry at Michigan
Technological University, Houghton.
RETHINKING THE THEORIES Enzymologists working on
their discipline's new leading edge need to make a "conceptual
shift."5 Enzyme systems do not reach equilibrium
even when they seem stationary; the system is in constant
flux. "Enzymologists have not paid enough attention to this,
but metabolic engineers emphasized the flux in biochemical
reactions for years," Qian notes. Characterizing enzymes and
their reactions in living biochemical networks, he argues,
means moving beyond usual equilibrium physical chemistry and
kinetics to include a branch of physics called nonequilibrium
thermodynamics. "This is the theoretical foundation of the new
enzymology," he says.
Traditional enzymology experiments aim to characterize, for
example, Vmax (maximum velocity) and Km,
to help define the enzyme's kinetic behavior. Km
offers a rough measure of how much substrate is needed to fuel
Vmax. But these parameters are often either unknown
or extrapolated from different organisms or species.
"Computing results from such a model is not going to be very
satisfying," Qian says. So, to produce such satisfying
results, researchers are examining a number of alternative
mathematical approaches, including stochastic modeling.
An enzyme molecule constantly collides with the surrounding
solvent molecules. As a result, the enzyme's thermodynamics
fluctuate rapidly. Traditionally, researchers regarded these
fluctuations as experimental noise, which could be ignored;
the bulk measurements of steady-state enzyme assays masked
such thermodynamic subtleties. But Qian notes in a recent
paper6 that such thermodynamic fluctuations could
contain valuable data, allowing enzyme kinetics to be modeled.
This means, however, shifting from the deterministic kinetics
of molecular concentrations in a traditional enzymatic assay
to a stochastic model, based on the probability that a
molecule will be at a particular state.
Traditional enzymatic assays are at steady state. The
researcher takes bulk measurements of the turnover of large
amounts of substrate molecules by multiple enzyme molecules.
But increasingly, enzymologists capture the kinetics of a
single enzyme molecule using a variety of methods, such as
confocal fluorescence microscopy and fluorescence correlation
spectroscopy.
So, enzymologists ask stochastic rather than deterministic
questions, such as, "What is the probability of the enzyme
being in an active configuration at a particular time?" A
stochastic approach illuminated, for example, ATPase kinetics
in motor protein movements. "Considering that the number of
enzyme molecules inside a cell can often be as low as a few
copies, there is a great need to think of enzymological
reactions as stochastic processes in the same way that
physiologists study membrane channels," adds Qian.
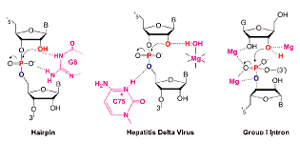
RIBOZYMES OPEN NEW VISTAS Meanwhile, scientists'
growing recognition of the importance of nonprotein enzymatic
actions has opened another new vista for enzymology. Victoria
DeRose, associate professor of chemistry at Texas A&M
University, College Station, and author of a recent review of
two decades' worth of research into ribozymes,6
remarks that enzymologists formulated ribozymes into
enzyme-substrate constructs that could be analyzed using
Michaelis-Menten kinetics. This allowed researchers to compare
directly the catalytic activity of ribozymes and protein
enzymes. "The power of enzymology as a discipline is apparent
in the way that ribozymes have been examined since the
discovery of catalytic RNA," says DeRose.
Researchers had "immediate interest in determining how, and
how well, RNA could perform the standard 'tricks' of enzymes,
such as positioning reactants and lowering transition state
barriers," she continues. "The new vista for enzymology is to
understand the mechanics by which RNA performs the roles of a
biological catalyst."
DeRose believes that electrostatics may be one of the most
important of the several structural differences between
ribozymes and proteins. "Internal electrostatics can play an
important role in catalysis, such as by altering the acidity
of a reactive group or neutralizing charges during a
reaction," she says. "RNA has a negatively charged
phosphodiester backbone with no natural positive charges,
requiring a counter-ion atmosphere. In comparison with
proteins, this is an exotic ionic environment."
Flexibility represents another important frontier in
ribozymology. A growing and compelling body of evidence,
including recent single-molecule studies, suggests that
structural changes influence ribozyme function.7,8
"It is important to determine the nature of these changes, and
to account for them in mechanisms based on kinetic
measurements," DeRose comments. "Ribozymes lend themselves
well to single-molecule fluorescence experiments because they
are relatively easy to modify and because they can have
large-scale movements that give clear fluorescence signals."
COMMERCIAL BENEFITS Meanwhile, some companies find
that advances in enzymology help hone their competitive edge.
"One of the most important advances in applied enzymology
recently has been the more general acceptance by the chemical
industry that enzymes can achieve things that are impossible
using traditional chemical catalytic methods," says
biotechnology professor Chris Bucke, University of
Westminster, London. (The cell factory discussed below offers
one example of this.)
Enzymes also provide a tempting and lucrative target for
pharmaceutical companies. "Most therapeutics mediate their
desired effect and unwanted side effects through enzymes,"
remarks Gunter Fischer, director of the Max Planck Research
Unit for Enzymology of Protein Folding, Weinbergweg, Germany.
Indeed, blockbuster drugs such as angiotensin-converting
enzyme (ACE) inhibitors, statins, and several AIDS drugs act
by inhibiting key enzymes. ACE inhibitors, for example, target
a key enzyme in the renin-angiotensin system, which plays a
central role in controlling blood pressure. Statins inhibit
the rate-limiting step in cholesterol biosynthesis (an enzyme
called HMG-CoA reductase), thereby reducing blood lipid
levels. Several AIDS drugs target key enzymes in the HIV life
cycle.
Researchers continue to target enzymes in their search for
more effective medications for common diseases. Approximately
200 million people contract malaria each year; perhaps two
million die because of it. The malaria parasite, Plasmodium
falciparum, increasingly resists conventional medications.
P. falciparum invades erythrocytes, destroys as much as
80% of the hemoglobin, and then feeds on the degraded
hemoglobin. Two enzymes (plasmepsins I and II) are critical in
driving the hemoglobin's destruction. Researchers have
identified several compounds that block these enzymes, raising
the prospect of new treatments.
The two plasmepsins belong to a large enzyme family called
aspartic peptidases. Other enzymes in this family have links
to several diseases, including renin (hypertension), cathepsin
D (breast cancer metastasis), b-secretase (Alzheimer disease), HIV-1
peptidase (AIDS) and aspartic peptidases secreted by candidal
infections.9 Drugs acting on aspartic peptidases,
such as the ACE inhibitors, are available now; others are in
development.
Insulin as well as many growth factors and cytokines act by
binding to specific receptors associated with enzymes, usually
a kinase. When the ligand binds, the kinase phosphorylates
certain amino acids that make up the receptor protein. This
autophosphorylation allows the receptor to interact with other
enzymes inside the cell, thereby amplifying the signal.
Mutated proteins in signaling pathways controlled by
kinase-linked receptors probably contribute to several
cancers. For example, signaling linked to the epidermal growth
factor receptor (EGFR) contributes to cell proliferation,
apoptosis, angiogenesis, and metastasis. Many drugs targeting
this and other kinase-linked signaling pathways are in
development or are marketed in some parts of the world.
Indeed, Iressa (gefitinib), which targets EGFR, is approved in
the United States for the third-line treatment of
non-small-cell lung cancer resistant to other
chemotherapies.10
Despite these successes, drug companies might not fully
exploit enzymology's potential. "To fully realize the
therapeutic potential of enzymes, a more knowledge-based
approach, rather than high-throughput screening, would be
desirable," Max Planck's Fischer says. "Enzymologists can
provide this knowledge by developing a more detailed theory of
how enzyme- ligand [interactions] and enzyme catalysis is
accomplished."
The pharmaceutical industry, Dandekar says, focuses too
much on in vitro or cellular systems, which may be a reduction
too far. "Often you get nasty surprises at the end of the
pharmacological pipeline from side-effects," he says. "We need
more top-down approaches, even if that is difficult."
Alabama's Johnson adds that sometimes the pharmaceutical
sector may jump the gun by targeting specific enzymes. "In
some cases, they try to develop specific inhibitors before it
is well established that enzyme X is actually involved in the
pathogenic process."
Enzymology also affects mundane aspects of daily life, and
there are few activities more forgettable than doing the
laundry. Subtilisin, which enzymatically degrades protein and
is used in laundry powders, is the most highly engineered
enzyme.11 Subtilisin must remain active at the high
temperatures inside washing machines. Campbell, who is also
president of the Nitrate Elimination Company, Lake Linden,
Mich., a biotech organization that produces nitrate reductase
and other enzymes for commercial purposes, notes that
comparing the structural characteristics of thermophilic
enzymes to those that function at lower temperatures may
reveal the properties that make the former thermally stable
and, therefore, more appropriate for commercial applications.
Westminster's Bucke, an applied enzymologist who divides
his time between industry and academia, adds that
site-directed mutagenesis improves industrial enzymes used to
make glucose and fructose syrups. "Native *-amylase, which is
used to start the hydrolysis of starch, is astonishingly
temperature-stable and is used at more than 100°C, but
requires calcium ions to retain that stability," he explains.
"Site-directed mutagenesis produced an equally stable enzyme
that no longer requires added calcium. This results in savings
in downstream processing."
The cell factory is another recent innovation that uses
genetically engineered organisms (such as Escherichia
coli) to synthesize particular proteins. There's nothing
new in that, of course. But enzymologists can engineer the
pathway used in the cell factory to make the protein and
eliminate unwanted reactions and control mechanisms. This
means that the organism produces higher protein yields with
fewer side-products than older genetically engineered
organisms. Bucke cites the example of a cell factory developed
by Genencor in Quebec, Tate & Lyle in London, and Dupont
that yields propan-1,3-diol, which is used to generate
propylene terephthalate at "cents per pound."
TACKLING THE FUTURE Elena Ghibaudi, research
assistant at the Department of Inorganic, Physical, and
Material Chemistry, University of Torino, Italy, argues that
enzymologists need to engage in improved dialogue and
information exchanges with researchers in other life sciences.
She notes that universities tend to train scientists to focus
their attention on specific aspects of a discipline; they
rarely offer a general picture. "People are trained to face
scientific problems from a relatively restricted perspective,"
says Ghibaudi.
On the one hand, this restricted perspective encourages
life scientists to develop specific competencies. On the
other, improved dialogue, information exchanges between
disciplines, and a cooperative approach could solve some
fundamental problems, says Ghibaudi. "Notwithstanding the
great number of conferences organized by life scientists all
over the world, I have the impression that not enough real
opportunities for discussion and confrontation between
disciplines are created." She calls for a multidisciplinary
approach, stimulated in part by systems biology. "By dealing
with cell complexity, systems biology implies
multidisciplinarity and a continuous confrontation between
viewpoints that are often far from each other," she says.
Speaking about complexity, Dandekar considers the discovery
of how enzymes work to be the most pressing problem facing
investigators. Four main research areas predominate: 1)
determining how enzymes achieve transition state catalysis; 2)
determining the protein sequence that leads to a particular
enzymatic activity or useful structure, known as reverse
folding; 3) characterizing how environmental factors and
allosteric interactions modify enzyme action; and 4) gaining a
better understanding of protein folding.
An additional challenge, says Fischer, lies in developing
reliable techniques for predicting protein- ligand
interactions or protein structures. For example, homology
modeling predicts protein structure from the primary amino
acid sequence, but the approach is restricted to proteins that
have similar codes to known structures. "We need a method to
predict structures and protein-ligand interactions ab
initio," he comments. Currently, poor understanding of the
basic physical and chemical principles and informatic
limitations hinder this approach.
For Michigan Tech's Campbell, "The most pressing question
in protein chemistry is: What are the rules for the 3-D
folding of a protein? ... [Researchers] need to know how to
design a new enzyme so that it folds to a stable structure;
then we can focus on active-site design so that new enzymes
can be created. We then need to understand how we can improve
the stability of existing enzymes to make more robust
catalysts that will be more useful in commercial applications.
Once we can design highly stable proteins with new and
improved catalytic activity, enzymology will be a mature
science."
All this without a razor blade in sight, save for the one
forged by 14th century philosopher William of Occam.
Mark Greener (biowriter@markgreener.fsnet.co.uk)
is a freelance bioscience writer in Cambridge, UK.
References
1. K.M. Donald et al.,
"Quantification of gene transcription and enzyme activity for
functionally important proteolytic enzymes during early
development in the Pacific oyster Crassostrea gigas," Comp
Biochem Physiol B Biochem Mol Biol, 136:383-92, November
2003.
2. A. Sharff, H. Jhoti, "High-throughput
crystallography to enhance drug discovery," Curr Opin Chem
Biol, 7:340-5, 2003.
3. H. Erlandsen et al.,
"Combining structural genomics and enzymology: completing the
picture in metabolic pathways and enzyme active sites,"
Curr Opin Struct Biol, 10:719-30, 2000.
4. S.
Liu et al., "Structural basis for the guanine
nucleotide-binding activity of tissue transglutaminase and its
regulation of transamidation activity," Proc Natl Acad
Sci, 99:2743-7, 2002.
5. H. Qian, E.L. Elson,
"Single-molecule enzymology: stochastic Michaelis-Menten
kinetics," Biophys Chem, 101-102:565-76, 2002.
6. V.J. DeRose, "Two decades of RNA catalysis,"
Chem Bio, 9:961-9, 2002. 7. J.H. Nagel, C.W. Pleij,
"Self-induced structural switches in RNA," Biochimie,
84:913-23, 2002.
8. J.E. Wedekind, D.B. McKay, "Crystal
structure of the leadzyme at 1.8 Å resolution: metal ion
binding and the implications for catalytic mechanism and allo
site ion regulation," Biochemistry, 42:9554-63, Aug.
19, 2003.
9. C. Dash et al., "Aspartic peptidase
inhibitors: implications in drug development," Crit Rev
Biochem Mol Biol, 38:89-119, 2003.
10. F.
Ciardiello et al., "Epidermal growth factor receptor tyrosine
kinase inhibitors in late stage clinical trials," Expert
Opin Emerg Drugs, 8:501-14, 2003.
11. W. Stemmer,
B. Holland, "Survival of the fittest molecule," Am Sci,
91:526-33, November-December 2003.
|