Does Evolution Have a Trajectory?
George Mobus
One of the first insights I ever thought I had gotten about evolution is that complexity has increased over time. This was at odds with much of conventional wisdom back in the 1960s. Biologists were loathe to characterize evolution as progressive because this smacked of teleology — that evolution was being pushed or pulled in a given direction, presumably by a supernatural power. The situation wasn't helped by the works of Teilhard de Chardin who, in spite of many valuable insights regarding evolution in general, insisted that the process was unfolding God's plan to create the noosphere.
Of course mainstream biologists gave no credence to this idea; so out with the bath water went the baby. It turns out that the noosphere is a worthwhile observation regarding organization regardless of what it may say about a supernatural power.
Today many biologists accept that evolution has followed a general pattern throughout the history of life on Earth. It remains mostly a naturalistic observation about the levels of organization in a hierarchy. This hierarchy involves increasing levels of complexity that have evolved over time. From the origin of life (or possibly even before) through the first simple, prokaryote cells, through the first eukaryotic cells and up to societies of humans, there is a recursive pattern of aggregating of cooperative individuals (molecules to people) and an eventual transformation of that aggregate into individuals that interact at a new, higher level of organization (complexity).
It would be hard to deny that this is a form of progression. Nevertheless, this new vision of evolution requires explanation that does not depend on teleology. Today, the phenomena of emergence and self-organization are being studied in many different systems contexts.
The Future of Evolution
I have spent no small amount of time thinking about evolution. And not just the evolution of life in the past. I also think a lot about the evolution of life in the future. In order to get to a point where thinking about evolution in the future makes sense one has to have discerned something of a pattern of evolution in the past. For much of the scientific history of the study of evolution Darwinian natural selection and chance mutations of genes (so-called neo-Darwinism) have dominated the generally accepted theories. The role of chance or random mutation played a heavy part in these theories as the source of novelty and variation upon which then, natural selection could operate to drive speciation. Jacques Monod's famous book, “Chance and Necessity” provided a general theory of evolution based exactly on these two factors. Monod coined the term teleonomy1 as an alternative to teleological, final cause, explanations for what appears to be a kind of ‘progression’ in evolution from the simple to the complex over evolutionary history.
In past blogs I have also written about the theories of Harold Morowitz regarding the flow of energy through local (materially isolated) systems and how that flow can drive a system toward higher organization. This perspective provides a reasonable causal, and purely mechanistic explanation for increasing organization in physical/chemical systems. The origins of life on Earth are couched in just such a theoretical framework.
In the systems book I am co-authoring I have been writing about the related phenomena of emergence (of organization) and something called self-organization. All of these phenomena now appear to give different perspectives on a universal process that generates increasing organization and levels of complexity so long as energy flows.
Figure 1, below, gives a sense of this progression. Starting at the bottom, under the lower dashed line, we have a materially-closed system comprised of many different kinds of components. The system can be thought of as starting in thermal equilibrium (and the components are stable, non-decomposable). The components will be well mixed given a sufficient amount of time passing in this state. The next level up shows some of the effects of energy flow, that is when an energy gradient is imposed and the system is driven from equilibrium. In this case we are mostly interested in components that combine in characteristic ways. That is, each component bears a unique ‘connectivity personality’ according to its type. Some of its potential for connecting with other components, either of the same or different types, is realized by absorbing energy from the inflow. As can be seen in the figure, some combinations are more complex than others. Not easily seen is that some combinations are more readily made than others due to the connectivity potentials across various types. Chance plays a large role in determining what combinations actually obtain initially. In the second panel we get a hint at what makes for a stronger association; the entity in the lower right side of the panel has a cyclical connectivity that helps stabilize the association. But local selection forces (i.e. the environment) will tend to break up some ‘weaker’ connections (third panel up). This makes some components available in the general pool and then the more complex, stable combinations can compete to obtain those to further increase their complexity, and possibly their stability (top panel).
A combination can be thought to be stable when the mutual linkages between components are strong; in a sense these components are cooperating to keep the bindings stable. This is in the face of selection pressures that might otherwise dissociate them. By chance, again, some more complex combinations are more capable when it comes to forming stable connections.
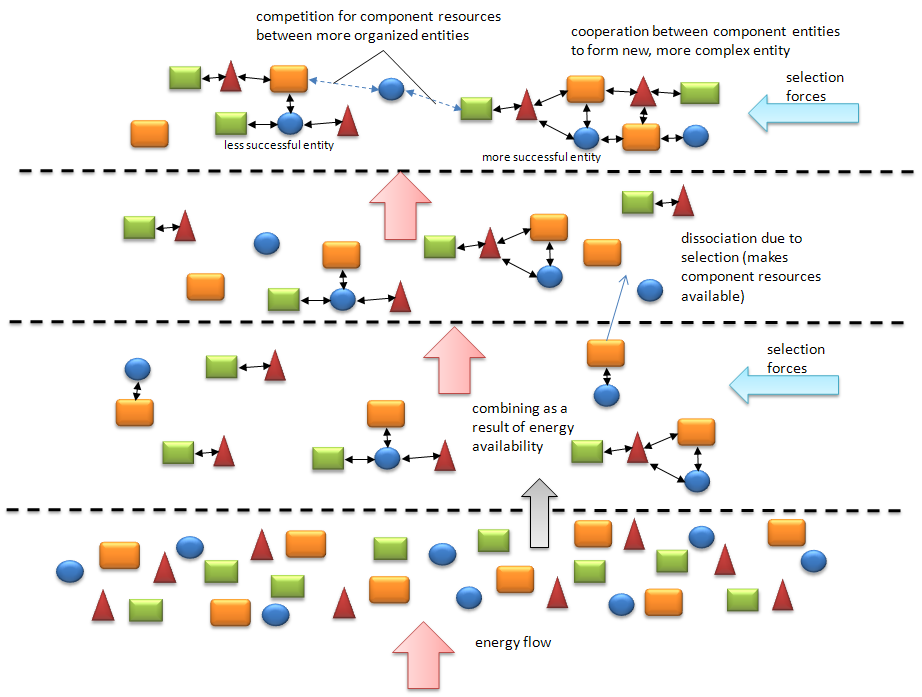
Figure 1. Evolution toward increasing complexity under the influence of energy flow. A materially closed system consisting of many simple but different components moves upward through successive stages of increasing complexity. Components combine in multiple, semi-random ways based on their individual “personalities” or propensities to bond (or be repelled). Some of these combinations are able to remain viable under the influence of a selective force (or forces) while others dissociate. As time goes on, with continuing energy availability, the more successful combinations, now acting as unified entities, are able to compete for remaining resources while others are not. Then, depending on the kind of selection forces encountered (as a result of this new environment) some of the more successful entities obtain resources at the expense of other less fit entities.
The above description of a pattern of increasing complexity can be seen most readily in the realm of chemical evolution such as occurred in the pre-biotic era on Earth. Much of this can be replicated in laboratory settings where starting with simple molecules in a closed container, and adding various forms of energetic inputs, the system produces a mixture of much more complex molecules, including some of the precursors of life. However, the above description is more general than just for chemical evolution. The nature of the components and the kinds of ‘personalities’ and connections that they make are different as we look at higher levels of organization but the pattern of progression is universal.
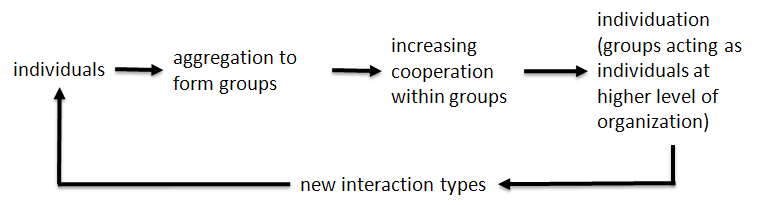
Figure 2. The process of individuation and subsequent group formation followed by the evolution of increasing cooperation within the group is a repeating cycle that has been seen in biological evolution at many different levels of organization. This shows the recursive algorithm-like nature of what is represented in Figure 1.
This pattern of progression, with transitions from simple to more complex, repeats itself throughout the biological complexity hierarchy (see below). There is a creative tension between cooperation between components forming groups or collections, and competition between different groups. Moreover, those groups represent new entities at the higher level of complexity. In the figure, the two ‘surviving’ complex combinations (struggling to acquire the unattached blue component) are unities (entities) in their own rights. As long as these combinations remain stable under the selective conditions imposed, and under conditions of excess energy availability, they will continue to sweep up free primary components as needed.
There is a limit to this process of incorporation of more components however. As combinations like this get more complex their energy requirements for growth and maintenance also grow. Under conditions of constant energy flow, however, these needs cannot be met and the entities approach an end to growth in complexity and size.
Worse still, from the competing entities' perspectives, if energy flow diminishes over time, then the process reverses and the combinations begin to loose complexity. If the environment changes in substantial ways, this too can cause the entities to fail, their structural and functional integrities to be lost.
The Origin of Life
The above description is surprisingly general. At first it seems to be slanted toward something like pre-biotic chemical interactions in the primordial soup that gave rise to life. And, indeed, it does cover that phenomenon. The necessary step to get from pre-biotic evolution to biological evolution is the need for entities to make copies of themselves rather than merely grow in size. Moreover, those copies must occasionally be subject to small errors that lead to slight differences in functions (behaviors) such that the selecting environment may bias further reproduction in the direction of one (or more) specific version. That biasing is because that version is better fitted to survive in that environment.
The step between mere increase in entity complexities, as above, and the beginning of what we might recognize as the first biological-like processes involves the beginning of cooperation between molecular entities. In Figure 1 I show two entities competing for a component. But it turns out that some complex entities (molecules) can actually cooperate with one another in a form of mutualism. These complex entities do not merely have form, they have functions, also determined by the flow of energy. Most of these entities are not inert, but take in resource components and produce product components under the right energetic conditions. Sometimes, those conditions are provided by nearby other entities that ‘assist’ in some ways. Those assistant entities may or may not get anything out of the process, but we can usually find the conditions in which both entities are rewarded for the interactions (in the case of an assistant that doesn't directly benefit, such as a catalyst, the benefit may come from a larger structural stability that keeps the assistant ‘safe’).
The situation is even clearer in cases where the product output of one entity is a resource input to another entity. And in cases where there is a chain of these interactions there is a possibility that the output of the final link can act as input to the first, i.e., they form a cycle. This will obtain when the chain acts to funnel energy flow through a sequence of work processes that ultimately give off waste heat at the end, along with components that will be available as inputs for the start of the sequence. These cycles need not be materially closed. They may form a stable structure due to some of the final outputs being needed as inputs to the start, but they may also operate on other inputs (for example smaller entities that contain usable energy) and produce usable products by other cycles. Figure 3A shows a schematic representation of cycle formed when four entities cooperate for their mutual benefits. For an example of a real molecular cycle see: The Citric Acid Cycle in Wikipedia.
Figure 3B shows another necessary step in the emergence of life, the development of an autocatalytic cycle. This is where an entity is capable of catalyzing the construction of a copy of itself from the energy/resource component inputs. Autocatalysis has been demonstrated in a number of molecular systems, including (and especially) in molecules of RNA.
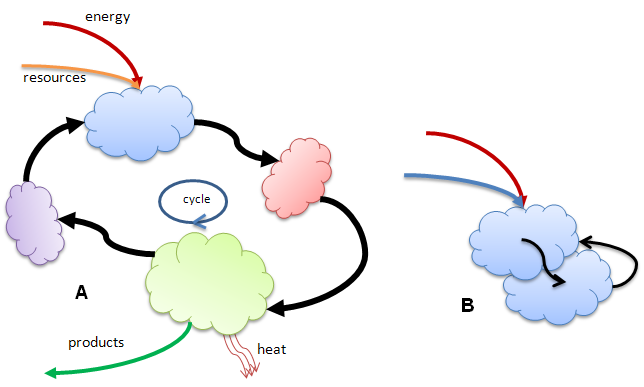
Figure 3. Cycles in which entities cooperate by using resource inputs and producing usable products in a chain that closes in on itself. Such chains are not strictly materially closed in that they may take in other resources, produced elsewhere, and produce products, not directly used by this cycle. A special kind of cycle occurs when an entity is capable of catalyzing its own generation. This is called autocatalysis and is the prime suspect precursor process for reproduction in living systems.
The key aspect of these cycles is the degree of coupling or connectedness involved to maintain the cycles over long periods of time. As a rule there needs to be some kind of topological constraint on the organization of the individual entities. For example a fatty acid membrane (which has a tendency to form naturally when clumps of fatty acids are immersed in water!) could contain these entities while allowing resources in and letting final products out.
The point is, of course, that these entities are not competing. They are cooperating, which is the source of higher-level organization and, hence, complexity.
My favorite molecule in the whole world is adenine! The reason is that it is a core molecule in many different chemical entities such as adenosine, the purine nucleoside that is in both DNA and RNA. Adenosine, in turn is a component of many more important molecular complexes such as adinosine triphosphate, the portable energy ‘batteries’ in cells, and in amino acid coupling. Adinosine does a spectacular job of linking with amino acids somewhat readily. In all of the various transfer RNA molecules (tRNA) adenosine is at the tail end of a cytosine-cytosine-adenosine chain that binds the amino acids. tRNA is the molecule that carries its amino acid rider to the ribosomes where the amino acids are unlinked from the tRNA and linked together to form polypeptides, the beginnings of proteins.
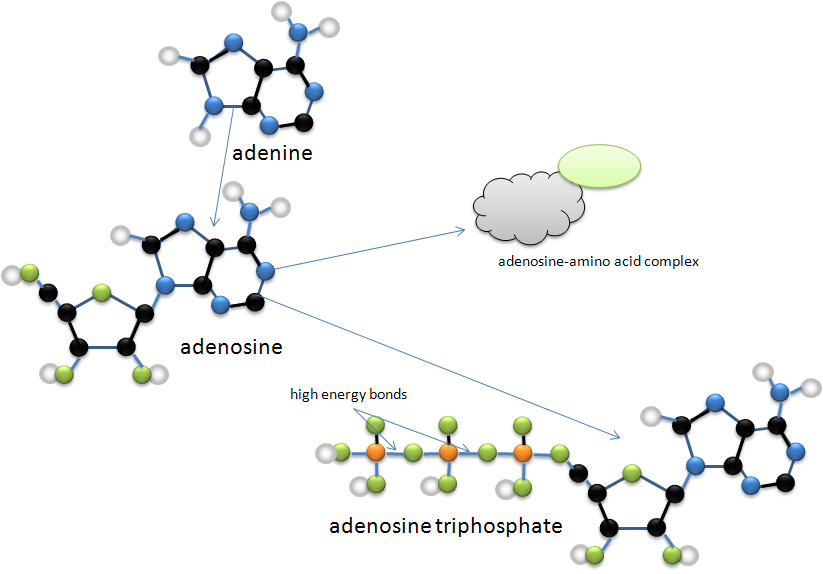
Figure 4. Adenine and some of its derivatives. In particular, adinosine, the nucleoside found in RNA and DNA molecules is multi-talented. Black balls are carbon atoms, blue ones are nitrogen, green ones are oxygen, white ones are hydrogen, and orange ones are phosphorus. Black lines are double covalent bonds, blue lines are single covalent bonds.
Adenosine is found in all of the major life chemical cycles in one form or another. My personal suspicion is that the origin of life will be found to involve the chemical complexities of this fascinating molecule that has so many functional and structural ‘personalities’.
Chemical evolution as described here is not really that different from biological evolution in terms of the basic concepts of competition (for resources) among entities, versus cooperation for mutual benefit. The former leads to the selection of the most fit entities within the constraints of the environment. This is just natural selection at the level of molecules. The latter, cooperation, is a natural consequence of functional processes accomplished by entities producing resources that other entities can use in a matrix of flows. So long as energy flows through the matrix, the flows of materials and especially the cycles of materials is sustainable. With the addition of autocatalysis (chemical reproduction) we have the basis for what makes biological evolution operate even more efficiently than chemical evolution. It does so because it achieves an optimal level of complexity that both provides for the creation and maintenance of structures and dissipates energy to prevent accumulation and cause heat death of the entities.
The Biological Hierarchy and Its Evolution
Cooperation among the components that make up an entity is the key to higher levels of complexity being achieved. We can make a good argument that a number of different cycles, both autocatalytic and simple, were jostled around in the primordial soup and smaller cycles found benefits from forming larger cycles. That is some form of early citric acid cycle coupled with some early form of protein synthesis mediated by RNA structures and RNAases (enzymatic RNA) coupled with some early form of RNA synthesis to produce the knowledge template upon which these cycles would be reproduced. Each cycle (entity) may have had its own successes in the environmental milieu of early Earth. But when they joined forces, the mutual benefits were spectacular. Life emerged from non-life. And thereafter, as long as the energy from Sol flowed through the Earth intermediate system, those combinations of primitive cycles and structures would continue to reach new variations in complexity that would be the beginning of early bacterial cells. From that point on, biological evolution, as a special case of the more universal recursive algorithm of cooperation discovery followed by the emergence of new levels of organization would govern further progress.
We've seen that at the level of atoms and molecules, the general rules for making connections (e.g. chemical bonds) and selection for successful combinations are relatively straightforwardly applied. But when living systems are involved, the mechanics are not quite as clear. Biological evolution involves surficially different mechanisms that might seem to hide the underlying universal rules. For example, the evolution of sex seems to complicate matters, almost giving a qualitatively different feel to biological evolution as compared with pre-biotic chemical evolution. There doesn't appear to be a chemical analog to sexual reproduction in the latter. Nevertheless, many biologists are now in agreement that the patterns of emergence, self-organization (especially in cycles), autocatalysis, and selection for the most fit structural/functional entities is the hallmark of biological evolution as well. Sex may be a special case of two different entities cooperating, and it just makes it more interesting ;^).
One of the major hurdles of late 20th century evolution theory was explaining various behaviors in social animal. In particular altruism, where an animal will potentially sacrifice its reproductive potential for the benefit of others in the social group seemed problematic given Darwin's version of evolution of more fit species depended on selfish competition. The breakthrough came when W. D. Hamilton (1963) proposed the concept of inclusive fitness in which both plain cooperation (mutual benefits) and altruistic cooperation (for the benefit of a shared genetic complement) could be explained in terms of selection for those behaviors when the underlying genes became more represented in the gene pool. Altruism requires that individuals have some level of relatedness so that the one that suffers a loss of representation in the next generation as a result of an altruistic deed more than makes up for it because the benefits bestowed on the recipients, who are related, ensures that more ‘related’ genes will be passed on. Thus the provider/sufferer will still have representation in the next generation even if he/she does not provide offspring directly.
The key here is seeing how some form of cooperation will increase the inclusive fitness of members of a species that drives them toward forming social groups with greater and greater cohesion — more intra-group cooperation than competition between members. The upshot of this dynamic is that groups that tend toward greater intra-group cooperation will tend to do better than groups with lower levels of cooperation if the groups are competing for resources in the environment (the species must have a high degree of niche dominance so that the species is out competing other species who may, say, eat the same foods).
This dynamic is exactly the same as we saw with molecular systems. A social group of animals that have high levels of cooperation between members, even when the members have many kinds of personalities, will tend to be more successful in the competition for resources.
The dynamic has been repeated many times in biological evolution. The first instance, demonstrating that it is not even necessary for the group to be formed from individuals in the same species (or related), was the mutualistic benefits that certain primitive bacteria and Cyanobacteria obtained by forming the kind of association depicted in Figure 3A. Lynn Margulis, in 1966 proposed that certain organelle in eukaryotic cells were actually remnants of previously free-living bacteria that had taken up residence in larger prokaryotic cells. She based this notion on the fact that some organelle in the cytoplasm of eukaryotes, such as the mitochondria, still have some of their own functional DNA, meaning that it specifies the construction of some parts of the mitochondria. This is a part of the genome which is separate from the nuclear DNA. The nucleus may have developed later from a prokaryote symbiont that had developed a more organized way of storing long strands of DNA in a package. It now appears that eukaryotic cells were the result of a tendency for many different species of bacteria to find mutual benefit by cooperating. A new kind of cell emerged as a result.
The story is told again when eukaryotic cells began to aggregate and cooperate for mutual benefit (actually many prokaryotic species also formed aggregates very early in life's history, but it was the eukaryotes that gave rise to what we recognize now as significant specialization between cell types). Both single species aggregations and multi-species forms have been documented. The origin of sex seems to have been when cells of the same species in an aggregate began to differentiate between germ cells and somatic cells. The former produce the gametes that recombine to form zygotes, new individuals. The somatic (body) cells begin to specialize in food getting and structure maintenance, etc. The differentiation into germ cells and somatic cells can actually be explained as a form of altruistic behavior on the part of somatic cells! These cells effectively give up their rights to produce offspring via meiosis and zygote formation. They are relegated to mitosis which results in eventual aging and senescence of the cell lines, while the germ cells get to participate in immortality! The phenomenon meets Hamilton's criteria for altruism since the multicellular form (the aggregate) benefits through increased fitness, as a unity, and both cell lines collectively enjoy increased inclusive fitness. The somatic cells are related to the germ cells, having the same genetic complement, so their genes are represented in the next generation of that aggregate.
Even when an aggregation from a single species of cells is involved, it is still possible that the genome of that species will be subject to revision by incorporating genes from other species. For example microbes that start out as parasitic on a larger multicellular species may eventually evolve to be mutualistic, that is, both species cooperate for mutual benefit. A good example is the microbes living in the rumen of cattle that can digest cellulose making the component carbohydrates available for digestion by the bovine. Both species benefit. But the story goes on. Under conditions of long term association (whether mutualism or not) we now know that genes from the microbes may be carried by various vectors into the nuclei and chromosomes of the host (larger or more complex) species. Thus the genome of a species may actually be an amalgam of the genomes of several completely unrelated species. Of course most of these errant genes will not be functional in the new environment. But occasionally they might be, or be subject to mutations that confer some functionality on them. As it turns out scientist now routinely perform genetic implantation of foreign genes in chromosomes for the purpose of causing the target species to produce a product or display a useful phenotype (e.g. tissues that emit or absorb light of a given frequency when activated!)
Some multicellular organisms eventually evolved the propensity to socialize. That is, at the level of a species population, these organisms formed aggregates in which individuals began to specialize or differentiate phenotypically and altruistically (as in the case of the evolution of somatic and germ cells). In extreme cases, such as the eusocial Hymenopterans (bees and ants, for example) this has led to distinct castes doing special jobs in the nest/colony. Some are workers, some are scouts, some are soldiers, and just a few are responsible for reproduction. Lest someone think this kind of organization is found only in the insect world, take note of the naked mole rats of Africa.
Less extreme (phenotypic polymorphism) forms of sociality exist. Many primates form social groups that have some degree of specialization and restrictions on reproduction. The latter is not always the case although one can argue that dominance orders serve to limit promiscuous reproduction, usually among the males. Some species of big cats (lion prides) have gone to polygamous organization where a single dominant male controls a breeding group. Other species (e.g. wolves and wild dogs in Africa) have a dominant breeding pair where the dominant female actually enforces the survivability rules (her pups get to live, all others must die!)
Humans have, as one might expect, the most interesting social organization of all! This is no chauvinistic bias. The human social organization seems to have elements from many different forms of social organization across many different kinds of species. We readily form groups based on cooperation with strangers. We each participate in multiple groups simultaneously. At the same time we feel perfectly comfortable competing with other groups. And we often find ourselves competing with others within our groups. We achieve this remarkable capacity with language. The linkages depicted in the figures above, in human aggregates, are achieved with communications. They are fluid, adaptable, and ephemeral. They are also subject to miscommunication. Yet we have succeeded at sociality in unimaginable ways and to unprecedented scales (think nations, even if these are only marginally successful groups).
The Human Social Hierarchy
The successes of human social organization is based on the same evolutionary mechanisms that allowed all other aggregations of cooperating entities to form new levels of complexity previously. Namely, large, complex human groups (societies) tend to organize in hierarchical fashion in order to manage the structure and processes more efficiently and effectively. As much as many of us might loathe the concept of hierarchy (as in a bureaucracy for example) for what is implied in a notion of top-down control, the fact is that this form is nature's way of ensuring that a complex system operates appropriately. And humans have tended to organize in this natural fashion.
What is wrong with human social hierarchies has more to do with the rapidity with which human societies have grown in size and complexity due to the inventions of culture and population expansion. Our social organizations are more like loose aggregations of individuals still being jostled about trying to find better fits. We are still in the trial and error phase of structure discovery and at the same time, are continually trying to find new relations within the otherwise natural order.
All of our various forms of governance, including economic management, are just experiments as the evolution of culture attempts to sort out what works from what doesn't. The fact that many of our experiments have not worked out so well should not provide a condemnation of the idea of hierarchical management. It only says that human sociality is still in an early phase of evolving.
Our sociality will evolve but it will do so embedded in the coevolution of culture and with the advent of powerful selection forces from the world we have been in the process of creating. A key ingredient in the advancement toward a more harmonious form of governance is the biological evolution of greater sapience, as I have covered in these pages often.
I imagine a future world in which energy-constrained human societies are much more circumscribed by physical realities. There will be numerous ‘tribes’ spread around over the habitable parts of the planet (after the effects of climate change play out?), and these groups will have better settled on the form of hierarchical management appropriately developed and scaled to their sizes and complexities. Such an organization could be as simple as the council of wise elders advising the members on all manner of social interactions. Or it could be as complex as a multi-tiered hierarchy with specialists taking roles in logistical and tactical, as well as strategic management, much like we see in mid-sized enterprises today.
The human case of social organization, and its evolution, is just the latest example of the universal rules of the emergence of levels of organization. Groups of humans are headed toward the same degree of within-group cooperation as has occurred in so many previous examples. Of course there are and will be differences in details. Humans are not dumb cells. And they are certainly not merely molecules. Nevertheless, we are enmeshed in the process of evolution with a trajectory toward the dominance of cooperation leading to the emergence of a new kind of individual entity based on groups of human descendants.
Our evolution is guided by the recursive rules of emergence.
The Rules
Darwinian evolution describes a process that gives rise to speciation by a simple set of rules. Replication with variation (errors or mutations in genotypes leading to variation in expressed phenotype and behaviors) followed by the environment's relentless selection of those variants most fit, those that can leave the most progeny. Neo-Darwinism merely adds a mechanical explanation for inheritance and variation by adding in the story of genes. And the observation of competitive behaviors add to the understanding of the mechanics of selection.
But as biologists have looked at the larger scale patterns of evolution, the nature of cooperation and the resulting drive toward cohesion and new levels of organization/complexity show us a grander picture of what evolution is accomplishing. As long as energy flow provides excess drive to keep shuffling components and be incorporated into linkages between components, evolution's arc is toward greater complexity.
What are the rules that produce this effect?
- Population of many different components (individuals) with multiple personalities:
There must exist a plethora of components, be they atoms or people. These components need to be of different kinds, each kind with different personalities in terms of what kinds of connections they can make between them. - Forces of attraction and repulsion:
The linkages or interactions are based on basic forces of attraction and repulsion. As components are jostled about due to energy flow, the various components have opportunities to interact and form linkages where mutual attraction serves, or propel the components apart in the case of repulsion. - Linkages involve flows:
The interactions between components involve the flows of material, energy, and messages (signals). Bindings between two components can be thought of as essentially zero rate flows. Messages are special cases of material/energy flows in which the flow rates/volumes are extremely low and depend on special transmitter and receiver/amplifier sub-components. Messages are time-modulated flows that can convey information to the receiving component. Flows may also involve intermediate buffers or reservoirs between the source and sink components in order to handle rate of production/consumption differences between sources and sinks. - Constraints within boundaries:
A necessary condition is that the system of components be bounded and constrained to interact within that boundary. The surface of the Earth is a gravity-bounded constraint that keeps components in an interaction zone (oceans, atmosphere, land surface, etc.) - Shuffling - energy flow:
Another necessary condition is that there is a flow of energy from a high potential source (e.g. the Sun) through the system (e.g. the Earth surface), to a sink of low potential (deep space). This energy does the jostling of components to keep them trying new interactions as well as supplying the energy to construct structures and drive processes. - Selecting conditions (including conditions created by combinations acting on each other):
The interior conditions of the system must test the combinations that are tried in the process of jostling and combining. The conditions cannot be so severe as to break every conceivable combination (e.g. the temperature cannot be so high as to break chemical bonds willy-nilly). - Some combinations provide inclusive fitness for the participants:
The testing conditions permit strong associations to persist, and be replicated, while eliminating competition from weaker associations in less fit combinations. - Emergence of a coordinator:
The final rule is another necessary condition that allows more complex combinations to persist. The more complex an aggregation of components becomes, the more opportunity for some components to adopt selfish behaviors, thus weakening the associations. As information chains (supply chains) increase in length there is a greater chance that some link will fail or degrade. This can be ameliorated by the advent of a coordinator component (a subset aggregate that functions to provide coordinating information, see the figure below). In the pre-biotic chemical evolution I contend the role of coordinator was played by adenine (as described above). After the advent of Darwinian evolution many different forms of coordinators can be discerned.
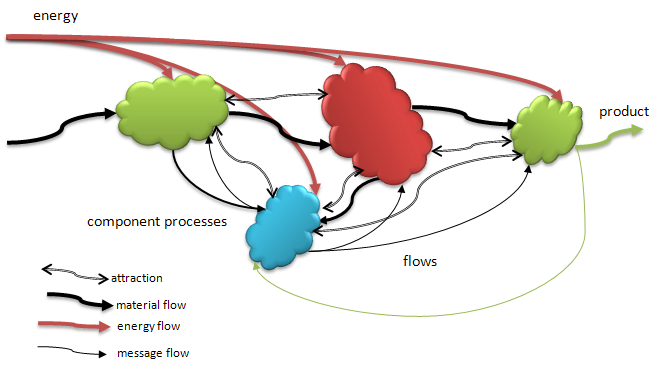
Figure 5. A complex organization of functional process components. These components are linked by some form of attraction or binding and have established flows of material and energy between them as they cooperate. The collective activities of this organization confers some survival advantage on the group that would not have been realized without the linking and cooperation. Note that the blue process is receiving various kinds of inputs from all of the other component processes and supply them with some ‘lesser’ inputs that it creates. Those lesser inputs are messages that provide information feedback more than resource materials. The blue process is in a position to act as a coordinator.
Figure 5 shows, in the general case, an aggregate of component processes that receive inputs of material and energy and produce products that are needed as resources by down-stream processes. The final process produces a product that is exported to the environment and is likely a factor in the success of this aggregate group in not being selected against, i.e. the environment finds the product useful in some sense. These components are bound to one another by some kind of attraction force that keeps them in proximity and allows the connections of outputs to inputs. The rates of flows between components is based on work done internally and mediated by the energy flows through those processes. Waste heat and material outflows are not shown but very much factors in the success of such aggregates.
Biological evolution adds a new twist to the cooperation within an aggregate of components like this. Once a means of providing for replication, with minor variations (e.g. mutations), of a successful aggregation is established, the processes are able to morph form and functions. If the change is minor and does not interfere with the primary success (function) of the aggregate, or even improves it, the Darwinian evolutionary rules apply and the system will begin to evolve through descent (with modification) and selection.
A special case of modification of function comes when one of the processes in a complex aggregate such as shown, takes on an increasingly important job of helping to regulate the production rates of the various members. A variety of means can be developed for doing so. But primarily the regulator-helper achieves its work and usefulness by receiving minimal material/energy flows from the other processes, extracts information from those flows, and produces feedback flows that engage those mechanisms that help regulate the behavior of the other processes. If the modifications to the presumptive regulator process (the blue one in the figure) are beneficial it will do a good job of balancing the flows through the whole system and, presumably, increase the fitness of the aggregate in its environment.
Note that at some point the component processes need not rely entirely on chance cooperation to maximize their inclusive fitness. The newly instantiated regulator provides information feedback that can modulate each process in order to achieve a more optimal result. This evolution of regulation assistance originated in a seemingly chance relation between component processes. In fact, the personalities of the components involved provided the right geometry for mutual attraction and binding that improved the chances of establishing the appropriate flow couplings and the final functional aggregate that could then enter the Darwinian process. Chance plays a role in jostling the components in random fashion, but that is just how the components get a chance to establish connections. There will be as many possible combinations as there are component types with potentially matching inputs/outputs. Bindings will obtain and some of those will result in functional coupling of flows. Others will not and the components will be unbound by entropic decay starting over the search for new relations. The bindings and couplings that result in beneficial aggregate behaviors will be preserved in the embedding environment allowing the aggregate to further replicate.
This development of a regulation helper that stabilizes or regularizes the behavior of a cooperating aggregate (system) is the beginning of the evolution of greater complexity in biology. It led to the origins of the first prokaryotic cells (with homeostatic maintenance and autopoiesis), to the aggregation and subsequent evolution of the first eukaryotic cells through the cooperation of various prokaryotic participants (endosymbiosis), to the aggregation of the first multicellular organisms, to the subsequent evolution of more complex organisms, to the evolution of groups of both conspecific (herds and colonies) and dissimilar (mutualisms) organisms, and to the evolution of ecological communities. All of these various groupings have different degrees or levels of internal cooperation and competition. Ecological communities display more instances of internal competition but still substantial mutualisms that benefit the entire community. The bodies of multicellular organisms are maintained by very high degrees of cooperation, but not entirely voluntary cooperation. Complex organisms and eusocial colonies are also characterized by regulatory mechanisms that constrain members to behavior that is mutually beneficial to all members. This regulation does not have to be viewed as necessarily coercive, but it fits the model of hierarchical control (see: The Science of Systems, 7). It is the latter that I now realize more clearly is the basis for the evolution of complexity in systems undergoing energy flow.
The Breakthrough — The Role of Hierarchical Control Structures and Their Evolution
I have spent no small amount of time trying to see how the hierarchical control structures, or cybernetic management systems, in nature fit the evolutionary model. These systems (actually subsystems that play the unique role of providing coordination to the operational processes) evolve out of simpler and somewhat ordinary operational precursors where the main inputs and main products are more informational than substantive with respect to the other processes (see Figure 5 above).
Pathways to and the Problems with Complexity
The role played by such regulation helpers is to keep the system of which they are a part operating smoothly and, as we will see, efficiently such that the whole system achieves higher fitness than those similar systems that lack internal coordination. The component processes of really complex systems with long internal supply chains cannot be guaranteed to behave optimally unless their inputs and outputs are monitored and balanced between all of the needs of all of the components.
The evolution of complexity involves the kind of specialization of subsystems within a larger system. The basic pattern seems to be that at some level of basic complexity the whole system achieves replication by each of the components themselves replicating and segregating to form newly bounded entities. Cell division is a case in point. It is possible that in the individual replication machinery a mistake occurs causing one subsystem to over replicate or form a redundant duplicate. If the mistake was repeatable then the new kind of entity has an extra copy of the subsystem that may or may not be functional. In any case the two copies of the redundant subsystem may now take on new trajectories in their subsequent replications. Errors (mutations) in the two copies are independent and can lead to different future functions in their lines. So imagine such a redundancy in copies of a regulation helper as depicted in Figure 5. Both copies replicate as required but in an occasional mutation event, one copy begins to develop additional information processing capabilities. A number of pathways might now open up to future generations of the system. In one pathway the regulator-helper that becomes more “sophisticated” in its regulation processing might compete with the original version and eventually take over the function completely but with much greater success in handling the coordination task. Or the same kind of redundancy error that gave rise to the regulator might affect other process components and they too might undergo further specialization needing increased coordination. Or the whole system might incorporate new, foreign components that provide new overall processing capabilities but that also need integration into the coordination matrix (remember the bacteria in the cow rumen?) There are so many ways this can go, but all ways lead to increases in complexity with concomitant needs for coordination regulation.
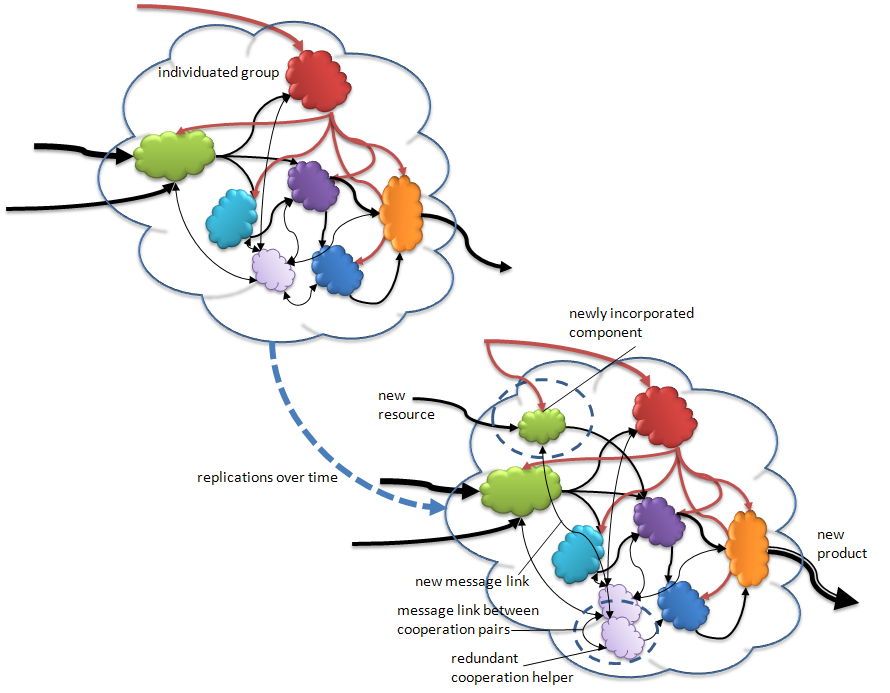
Figure 6. Over time complexity increases through evolutionary processes. The original group that has undergone individuation (upper version) replicates over many generations (to lower version). In some generations of the replication, errors (mutations) lead to various variants. See text for explanation.
In the above figure we see some possible pathways to complexification in the individuals in a population. During these replications various changes can occur. In the lower version in the figure we see variations resulting from the accidental redundant replication of, in this case, a coordination helper component (double headed thin arrows represent message flows between components). We also see that a new (foreign) component has inserted itself into the matrix, as in endosymbiosis, and managed to get linked into material and message flows to begin the process of integration. We also see that this results in changes in the final product produced. If the new product puts the individual in a more fit postilion relative to the environment it may then out compete its sister individuals for resources and energy; it can then go on to out replicate the competition.
Individuation includes the specialization of components. In the above figure I show (in the upper version) a single resource capture component and a single energy capture component. Both of these are then required to distribute their gains to the other components so that they can transform them into useful products (which, though not shown, includes replicates to form a future individual). I show a single final product assembly component. Another pathway to complexity is for any or all of the components to replicate without being expelled in a new individual replicate. After that time chance replication accidents in yet future generations may cause the redundant components to further specialize or even take on whole new functions derived from their original form.
At some point a new phenomenon entered into the complexity story. As a result of basic complexity continuing to evolve (i.e., more components and better coordination) organisms began to develop more flexibility in their response capabilities to environmental changes. Rather than achieving some kind of long term fitness because the environment in which they lived stayed stable for long periods, organisms became much more adaptable by having redundant systems that could take on different roles on demand. The advent of adaptive response mechanisms allowed organisms to do a much better job of internal coordination as well as external interaction management with objects in their environments (tactical management). An adaptive response mechanism is one that has latent potential to deal with variations in the demands placed on the organism in a wider range of habitats. For example the capacity for muscle tissues to increase in bulk if the organism finds itself in a more physically demanding environment (e.g. fishes that invaded areas where tidal flows were stronger than they had been used to). Humans undergo this kind of adaptation to work loads (or lack thereof in our modern mechanized world). The key understanding in adaptive response mechanisms is that resources will only be marshaled to meet the demand and otherwise be conserved or routed to other mechanisms with other demands. Internally the organism's coordination subsystem incorporates a ‘budget’ process for managing resources between potentially competing subsystem components. Again, evolution would favor those systems that evolved more efficacious management budgets, especially those that reacted quickly to crises situations (see my paper: Foraging Search — Prototypical Intelligence for a description of adaptive response, especially as it pertains to synaptic plasticity and learning).
Adaptive response actually evolved very early in living systems (prokaryotes and eukaryotes alike) but really took off in multicellular organisms. But it created a new phenomenon that would, forgive the pun, complicate matters. Having multiple component processes (subsystems) that have a much higher range of response capabilities means that more coordination is required, indeed a very sophisticated level of coordination (that resource budget). It also means that organisms, especially non-sessile animals, have something of a matter of choice in selecting appropriate behaviors. At first the choices amounted to entering or leaving an environmental situation (perceiving the situation followed by an action that depended on the potential consequences of the situation). To make those choices required much more sophisticated sensing and motor control capabilities as well as some kind of decision processing ability. Thus was born intelligence as we have come to think of it. Evolution of coordinators had now reached the level of brain systems and had led, in turn, to many more evolutionary possibilities.
Both the further evolution of sensors, actuators (operators), and decision processors, as well as the real-time decision making of individuals were coupled in an arms race with the rest of nature. Nature itself changed as a result of changed behaviors of species, populations, and individuals therein. In each generation new possibilities arose. This meant new problems had to be solved, either by selection of new genetic variants or by modified behaviors by individuals. Either way the end result is increasing complexity of the cybernetic system and concomitant increases in complexity, if not raw abilities, of the other machinery involved in coordination of the individual with the environment.
Human societies are also capable of adaptive response. In combinatorial explosion! Joseph Tainter recognized this pattern of complexity increase reflexively causing new problems to obtain, causing a need to find new combinations of facilities to respond to the problems. And that simply leads to new complexity. The cycle continues. In the realm of biological evolution the cycles are long and drawn out but observable in the fossil record (and the genome record). But in human societies the cycles are rapid and growing more so as time goes on. We see the results in the nature of our technologies and our social institutions like governments, enterprises, and even social clubs. They become complex in cultural evolutionary terms and that complexity creates new problems as the institutional entity engages in a now more complex world. But Tainter also saw an interesting phenomenon taking place in societies that had undergone significant complexification over their history. Complexity seems to have a point at which the benefits of solving problems begins to diminish with increases. In other words, solving the next problem by increasing the complexity of an already complex institution does not help achieve a stable situation and only leads to more problems. At some point further, increasing complexity, which is the natural response of the system, actually carries costs greater than the benefits expected. But because the only response the system knows how to make to increasing problems (and their costs) is to go on increasing complexity. In Tainter's view this is the major cause of collapse of historical civilizations and is likely operative in our current developed world situation.
The phenomenon seems real enough and we can see it in the natural world, though not as clearly as in the human world. Many more authors have been agreeing with Tainter's thesis. A nuance to the thesis, one that Tainter has recently been paying more attention to, is that the ability for complexity to be supported is based on the flow of energy (and other resources) through (and into) the system. Most of the civilization collapses that he and others have studied showed signs of diminishment of energy flows followed collapse of the institutions that were maintained in complex working status by the former levels of energy. Thus, we see that complexity is actually driven upward as a function of energy abundance enabling adaptive responses to changing environments. This generally works out for biological evolution. But the problem for humans is that the time scale of cultural evolution is grossly mismatched with that of biological evolution. Therein lies a fundamental problem for complexity management.
Nature's response to increasing complexity is the evolution of hierarchical control structures fitted to manage that complexity. In animals we see this in the evolution of increasingly capable brains able to deal with the coevolving complexities of the environment. In human societies we see a very rough approximate attempt toward having functional hierarchical control systems develop that could coordinate the underlying complexities of institutions. But the pace of change in human societies is simply too great for evolutionary processes to keep up. And by that, I mean the biological nature of the principle components of societies, humans, has not kept apace with the rising complexity such that human-based coordination structures (those coordination helpers from above) could evolve to handle and manage things adequately. Most of our governance mechanisms developed over human history, at least since the advent of agriculture, have had their problems and failings, mostly because they are built using human components (though those same humans striven mightily to find systems of laws and procedures that could compensate for human failings, with some successes, but mostly failures — laws are fine but people cannot be relied upon to obey them!). The most modern example of this is the failures of democracy (i.e. the representative kind supposedly practiced in the United States). The system to be governed is simply too large and too complex for human designed government subsystems to handle effectively. Think of the arguments over strict constructionist interpretations of the Constitution of the US and the problems arising due to technologies for which the founding fathers could not have anticipated the consequences. Even corporate governance is apparently badly broken. Enron might be the poster child for that problem.
Human beings, as currently constituted, simply make bad components from which to construct an adequate management hierarchy for the level of complexity we have created in our world by being so mechanistically clever. From my perspective this is part of the weak sapience problem (see my sapience thesis: here). Part of sapience involves the build up over a lifetime of functional and veridical tacit knowledge about how the world works which provides the basis for wise judgments guiding decision making. The current species of humans is lacking in the level of sapience needed to form effective management hierarchies fulfilling the pattern of hierarchical control as nature uses. The reason is simple and, in some sense, fixable. Humans, as components in groups, are still very oriented toward competition. This is aggravated by the fact that the kinds of groups that humans tend to associate with are not the ancient tribal (extended familial) groups that we evolved to live within. Rather, we humans associate in loose aggregations with cooperation employed when it seems best to accomplish a common goal. At many other times competition dominates our modus operandi even between group members. We are as often as not unable to even understand what our supposed shared goals are. Humans are independent thinkers who will pursue their own agendas unless they see a clear personal benefit from cooperation. Notably this is far more pronounced in western cultures where, in the extreme, rugged individualism is often lauded as the ideal human condition (of course just don't mess with my medicare!). Socialism is branded as a bad thing because it diminishes personal freedoms. And in America (probably more than anywhere else) individual freedom is held in highest regard.
In eastern cultures, especially Asian cultures, there is a greater sense of the collective as having importance, sometimes even above the individual. Westerners will sometimes denigrate this tendency as too insect hive-like. Remarkably, however, westerners fail to see their own condition is functionally no better than easterners. We in the OECD countries, for all our praise of individual freedoms are just as much slaves to our jobs (if we have one) in general. As Pink Floyd famously intoned we are “...just another brick in the wall.” That condition enabled by our education systems!
Nevertheless, western ideologues insist that personal freedom is the good and the collective (mind) is the bad. They are trying desperately to hold onto an ideal that may have helped humans survive while they were invading new territories but has become dysfunctional in the modern crowded urban world. The reason these people cling to their cherished beliefs in individuality is that they cannot imagine any other model for a collective mind that doesn't look like the insect (eusocial) models where each individual is just a cog in a wheel and effectively sacrifices their own well-being for the benefit of the hive. This is really just a failure of imagination and an understanding of what cooperation and coordination in human societies might be based on. Our only experiences with experiments in coordination (governance and management) seem to all be ending badly because of all our human failings as ordinary Homo sapiens.
Humans need to evolve greater sapience. Effectively they need to become super sapient or truly sapient (what I call Homo eusapiens). With functional wisdom will come a capacity to make good, morally justifiable judgments about the logistical, tactical, and strategic management of our societies. Without functional wisdom we make egregious errors (think Alan Greenspan at the Fed, or Obama now!) and worse yet, never really learn from those errors (part of gaining greater wisdom for the future). This is precisely what we see happening in our civilization today. We have reached the point of diminishing returns (indeed probably negative returns) on complexity because we have not created the appropriate management structures composed of competent management components. Eusapient people will not be any less individual in terms of their capacity to think new thoughts. But they will have a better basis for thinking valuable new thoughts that will benefit the whole society. They will have a much less selfish (profit motivated) attitude toward their own position in organizations. I suspect that the Asian tendency toward greater interpersonal cooperation and even what appears as submission to authority are remnants of Homo sapiens emergent capacity to form cooperating groups that evolved prior to agriculture and persisted to some extent with cultural evolutionary turns that tended to preserve older customs (they also tend to value wisdom of the elders more so than westerners do). In the western and middle eastern world cultures became too complex too quickly and conditions for the dominance of cooperation were largely subdued. Then the evolution of hierarchical coordination mechanisms (governments), nature's own way, was corrupted by the fact that the components of the coordination subsystems were simply not, themselves, sufficiently strong cooperators. They were not yet sapient enough.
Managing Complexity
To be fair, it may be the case that we are simply over extended in complexity. It was made possible by the wealth of abundant high-power energy in fossil fuels that allowed us to stumble upward in complex institution building, always compensating for our bumbling creation of new problems by building new layers onto our already inadequate management infrastructures. Think of all the laws we have to create and then revise and then repeal and then rewrite and renew as we create an impossible governance infrastructure. Think of the tax codes in the US! How more convoluted can you get? Think of the subsidies for corn ethanol to solve the high gas prices. It simply contributed to higher food prices as corn was siphoned off the food chain and ended up in our gas tanks. Think of the congressional mandate to blend gasoline with 10% ethanol. What problem did we actually solve? How many more problems did we create?
How does the problem of managing increasing complexity get solved in nature and biological evolution? And what might be the consequences for human societies of imposing nature's solution on our human condition? I will tackle the first question in this section and the second question in the last section below.
The study of hierarchical control systems in nature reveal some repeating patterns that suggest a way that human societies might become successful as groups with common beneficial outcomes.
The first lesson comes from the way in which components couple in successful aggregations (as described above). Strong attractions bring components together and linkages for mutually beneficial flows establish productive functions. In humans we do feel attractions toward other individuals and groups. Whenever we get excited about entering a relationship (of the non-romantic sort) we are experiencing nature's tendances to bring components together and form basic bonds. Then if the other produces some behavior, even just the kinds of communications that stimulate our mental states, or some other production that we find rewarding we find our coupling strengthening. We develop the emotions of liking the people we associate with.
On the other hand, if the original attraction isn't followed by satisfying flows of resources (communications especially) we can develop repulsions, or certainly weakening of the original bonds such that we are susceptible to attractions from other individuals or groups. Our human nature is to seek associations and we have to first see if there is something attractive about those potential associations before we move. In the same way we are often repulsed by groups or individuals from the start and would never seek relations with them — we continue our search for attractive associations.
What makes an association turn into a solid group process is success in producing something that everyone values. And those values (within the group) derive from the value the embedding society (the environment) places on the product. In other words natural selection is at work to select for groups that work well as a unified entity. The bindings and regulation of flows of resources through the group are initially loose but are mediated primarily by feedback communications between flow partners. The receiving partner lets the sending partner know what s/he is expecting, or alternatively the sending partner can let the receiver know what s/he has to offer. It is this mutual partner-to-partner communication that is found most prominently in small groups where all of the component members are in close communications with everyone else.
At some point groups of this kind, what I would call a production group, become concerned with maintaining the quality of their products. Perhaps environmental feedback has made such attention necessary. The first form of cybernetic regulation of a process is that of simple feedback. The quality/quantity of a product is sensed by some specialist member of the group who reports the results to another specialist who, in turn can evaluate the results and send a message to the producers to get them to change their procedures if necessary. Operational control evolves as a means of maintaining the fitness of the process (of the group) in simple systems. Figure 6, below, shows this simple version of feedback control-based management. Note that we are assuming that the internal flows and attractions are maintained by the members of the group. The producers must recognize the evaluative feedback as valuable and not a kind of mean criticism. At times even this simple process might be jeopardized by low sapient minds!
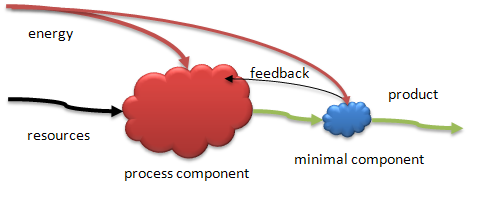
Figure 6. The beginning of cybernetic (control) systems. One component processes resource inputs, given an energy source, and produces a product that is detected by another component that doesn't change the product but, itself, produces a message that feeds back to the producer. See text for details.
As groups evolve and become more complicated a more complex form of feedback control is needed to coordinate the whole. This is shown in Figure 7 below. In more complex systems, each component is responsible for some intermediate function in contributing to the final product. Here we show an increasing chain of intermediate processes. In a situation where there are simply many identical parallel processes producing the same basic product the control system is still as simple as in Figure 6, but with as many feedback loops as needed to modulate each individual worker. Figure 7 depicts something like an assembly line where the final producer does not know or see what the first producer is doing. Here the problem is that the close coupling communications between all members is not feasible and that creates the need for a specialist that can act as an information distributor. This is the earliest form of logistical control, the balancing of the needs of all the component process and making sure the resources are optimally managed through the process. It is one step up in the control hierarchy.
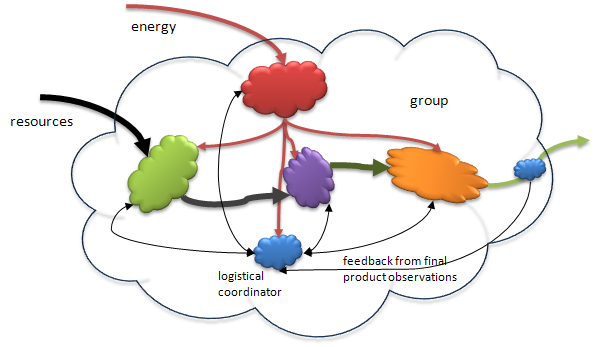
Figure 7. In more complex groups producing a product, the same basic feedback system has evolved so that intermediate products in a supply chain (assembly line) can be monitored and information fed back to all of the components involved. Each of the components report to the coordinator (the blue blob at the bottom of the figure) as to the quality/quantity of the intermediate products coming from the previous component (green to purple to orange). The coordinator then sends messages to the components earlier in the chain to have them make adjustments (if needed) so that the whole process is optimized. The final, small blue blob, is the same as the detector shown in Figure 6. It provides feedback regarding the quality/quantity of the final product. .
In this system the component that is responsible for acquiring the resources and the component responsible for acquiring the energy (or as I have pointed out in other writings, the equivalent in monetary resources) are both assumed to be passive receivers of their respective inputs. As systems become a little more complex they acquire the ability to actively obtain resources and energy. Animals eat. Humans create purchasing systems and accounts receivable (or taxes)! They may also develop the capacity to manipulate their products, sending them to other entities as opposed to simply passively putting them into the environment (note that such behavior is flow coupling!) The input and output processors become active with respect to the environment and their activities must now be coordinated not just with internal processes but with other entities and processes in the environment. The interactions of the entity (individuating group) are not usually predictable and may vary in terms of which other entities are involved or over time. In other words, the control of the entire group now includes tactical management, not just logistical and operational control.
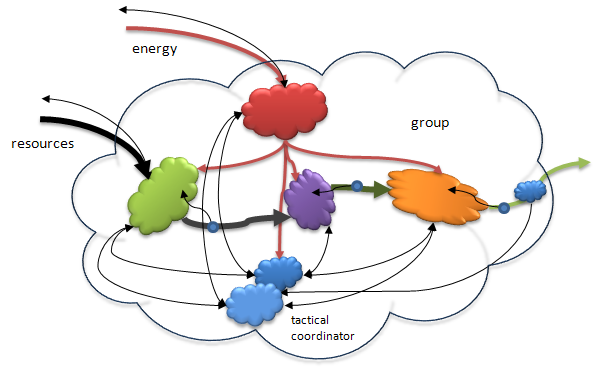
Figure 8. When the resource/energy acquisition and product distribution components evolve to become active agents a new kind of coordination control is needed. Here we see both logistical (internal coordination) and tactical (external coordination) controls. These two coordination controllers must also cooperate so that the tactical controller is issuing instructions to get the needed inputs in order for the logistical controller to produce optimal outputs. In this figure I show that the various internal (intermediate) components have evolved their own versions of operational control by monitoring their own outputs and using the information to modulate their own activities. This figure then shows the rough relationship between the two lowest levels of hierarchical control, operational and coordination.
The controller components thus far need to be complex enough to form representations of models of the component process that they affect. The logistical controller must have a model of the whole production process that allows for determining what an optimal operation entails. It uses this model to send adjusting signals to each of the components as needed to achieve the final optimal output. Similarly, the tactical controller has a model of both the operations of the active acquisition and disposition agents as well as a basic model of the external environment insofar as it can predict that environment's behavior, which might affect the need for changes in the activities of the agents. Both the logistical and tactical controllers require a good deal more sophistication in terms of their internal information processing capabilities. Animal brains reflect this same pattern of distribution of responsibilities and processing capacity. Basic operational controls range from reflexes to monitoring and regulating breathing or heart beats. Logistical and basic tactical controls are realized in so-called limbic (midbrain) areas. These include things like making decisions to increase the heart rate and issuing orders to the operational nucleus that activates the response. The tactical controller (e.g., the amygdala) may have sensed a threat that required increased respiration levels in the body to escape.
Tactical control requires not just a fixed model but a model that includes memory for storing variable states of the external world that is relevant to the entity. In simple human organizations that can be supplied by human memories. In more complex, sophisticated organizations we rely on written or computer records and computer programs to manipulate the data. For example a database of sales can be mined for important information about the behaviors of customers. That information can help predict demand and future sales. In animal brain evolution the limbic system evolved a primitive cortical mechanism and more sophisticated long-term learning to construct conceptual models used to determine what elements of the environment were going to do in the near future. A simple example is the mental processes needed by a carnivore when it is chasing its prey. It needs a basic model of what the prey's behavior might include and a memory of the most recent trajectories of escape to predict where to direct its efforts for a successful kill. Dinosaurs (and modern day lizards and alligators) didn't need elaborate brains, just enough cortical tissues to handle the behaviors of their, presumably less sophisticated, prey.
The final (highest) level of control in the hierarchical model is strategic management. This capability evolves from the need to extend the time and space horizons beyond the relatively immediate ranges. At a sufficient level of complexity of the environment the organization (brain) needs to not only know what the entities that it immediately interacts with are doing, it also needs to know what the entities that those interact with are doing in order to do a much better job of anticipating the more distant future. If the anticipated future includes possible major changes to external conditions then it may also be necessary to reorganize the internal structures of production to take advantage of future opportunities or mitigate future threats. Reorganization of the internals is handled by a more sophisticated logistical control system. Birds building nests in strategic locations and humans considering producing a new product are examples of adaptive response extended in space and time to maximize fitness while keeping the organization alive and intact.
The human brain is the best example we have of a most sophisticated hierarchical control system functioning within an extremely complex biological system. We are examples of multicellular aggregates that have achieved exemplary cooperation through the hierarchy of coordination mediated by tight coupling and intricate communications between all of the components. Every cell type in the body has a specialization and does its job for the good of the whole. Yet there is an aspect to these specialized cells that has only recently been revealed, that I think may pertain to the human condition. It had been thought that even though every cell (except red blood cells that have enucleated) retains a complete complement of the genes passed from the parents through the germ lines, once cells had differentiated beyond a certain point they were stuck as those specialists until they died. Ordinarily this is the case. The only exception is when a cell turns cancerous and breaks out of its role to become malignant, essentially reverting to an undifferentiated state capable of endless division (until it kills itself by killing the person). But we now know that some kinds of specialized cells can be manipulated to cause them to turn into pluripotent stem cells. It is a matter of unlocking the silenced genes that are responsible for this state. The point I want to make is that in spite of the process of differentiation causing cells to become specialist and cogs in the really big wheel each retains the capacity to be an individual, like a zygote. It is all a matter of environment and needs.
The Significance for the Future Evolution of Human Society
The reason I mention this fact about cell dedifferentiation is that the same principle applies to humans who have become specialized for the good of society. We actually do sanction this specialization by virtue of how we educate people today. The operative meme in our society has become that education is all about getting good jobs (in high paying fields hopefully). This puts an undue emphasis on specialization that carries all the way through to the production of PhDs in various fields who will turn around and “train” new workers in that field. In order to get tenure you have to prove beyond a shadow of a doubt that you are a good specialist and that milieu perpetuates the meme.
In truth the human brain remains relatively malleable (sort of pluripotent) well into adulthood and in some cases even late life. Our capacity for learning is immense and should not be channeled so heavy handedly to specific silos of knowledge. My own suspicion is that this system of emphasis on specialization is driven by a very simple fact. The average human is just not sapient enough to achieve viable pluripotence throughout life. By this I mean that the capacity to use whole systems thinking (see my work on sapience to see the role of systems thinking in sapience) to grasp the essence of all systems of knowledge is not sufficiently well developed to allow most people to feel comfortable in multiple disciplinary fields. Systems thinking provides a basic universal understanding of knowledge that is transferable to any field. Very few people have this ability. Moreover, the sheer memory capacity needed for extensive tacit knowledge as well as adequate general intelligence (e.g. the capacity to form and recall memory engrams) to capture and integrate that knowledge is much more than the average human possess. Greater sapience would provide people with a much greater capacity to be specialists in one area and then learn to be specialists in another as the needs of the society and their own personal interests require. In other words, sapient people, exercising the maximum of cooperation and altruistic behavior — happily being part of the collective mind — need not get stuck just being a cog in the wheel. Sapient humans represent the ultimate (so far as we know) of adaptive response mechanisms!
From nature we have extraordinary examples of hierarchical control systems, their architectures, their functions, their performances. The human brain represents the most sophisticated and capable such structure in an individuated group, the human body. We could learn a lot about coordination control from its example.
It is actually fairly easy to imagine what kinds of governance (both social and economic) structures would obtain from this model. We actually have poor attempts at such institutions now. We were in the process of evolving these as nature dictates. But our hierarchical systems are poor when it comes to managing the complexity of our societies. They are failing, not because the basic architectures are wrong, but because the components out of which they are built are faulty or substandard based on the needs.
Imagine a governing body consisting of eusapient beings. What would it look like? For one thing I'm betting it would be a lot simpler than our current US federal government system. Why? Because it would not have to be constructed of layer on layer of complex mechanisms meant to compensate for the failings of the next lower layers. For example if enterprises were run by and comprised of more sapient beings they would not need an EPA to monitor their waste outputs and assess fines on them to regulate their behavior. Firms would do the right things at the operational level. Market economies would be so much more able to coordinate trade. Even the idea of a planned economy (logistical control) would not be the pariah concept it is in capitalistic systems today. The problem isn't that planning doesn't work. The problem is that it has always been poorly executed by much less than adequately sapient beings!
Interested readers can read my working papers on sapient governance here.
What is Required
This is the difficulty. In order for humans to have a sapient society they must, themselves be sapient. Or rather, as I said above, they need to be super sapient. But this is no surprise. We are in the midst of a coevolution between human biology, specifically the brain, and cultural systems of governance. What is needed is for humans to evolve more sapient brains in order to be better components in the highest organization level that has been evolving on this planet. In order for human groupings, e.g. manufacturing companies, or farming collectives, to achieve individuation sufficient to become truly effective and efficient entities humans will need to develop eusapience.
How this is to be achieved unfortunately involves a new speciation within the genus Homo and (this is the hard part) the extinction of our current species. Now this need not be a completely traumatic process were it not for the sheer massive numbers of humans alive on the planet today. There are plenty of examples of sympatric speciation (a sort of transition to a new species with temporal, inter-generational separation reinforcing behavioral differentiation) having taken place. But we have, unfortunately, over populated our habitat and this will lead to a more traumatic transition, within just a few generations. I've written sufficiently about this eventuality so don't want to repeat myself here.
Suffice it to say that I think Joseph Tainter's notion of collapse will ensue, but on a planetary scale. That means the direct survivors of collapse will have no place else to escape to. The complexity of human societies will be severely diminished in a very short period of time. What I believe must eventually come to pass is that a new variety of humans, those that have sufficient wisdom to survive the future world, will emerge to form new and better groups. The pattern of individuals forming groups and groups evolving through individuation to new entities at a new level of organization will reestablish. It will be made possible because humans will catch up in their biological evolution to allow for strong stable interactions just as the molecules and prokaryotes and eukaryotes and tribes did before them. It is the grand trajectory of evolution so long as energy flows through the system.
1 The Wikipedia article is marked as being disputed. After reading both the article and the associated ‘talk’ page I am not sure what the point of the dispute is, other than some quibbles about the article giving undue bias to biological evolution when in philosophy the subject is broader and considered ‘technical’. I find the article suitable for at least a preliminary background. My treatment, in the text above, of the role of energy flow in driving processes should make clear that teleonomic ‘pseudo-purpose’ is legitimately different from teleology.