Human Island Biogeography
This page examines some modeling issues in island biogeography, particularly as it applies to humans.† From this page, you can look at some of the modeling ideas our group has been examining.† We present some highly simplified models that try to capture the most important relationships between human populations, populations of different prey species, and island ecosystems.† Our goal is to develop these models into a framework that, combined with empirical observations, will further our understanding of the interactions of humans and their environment.
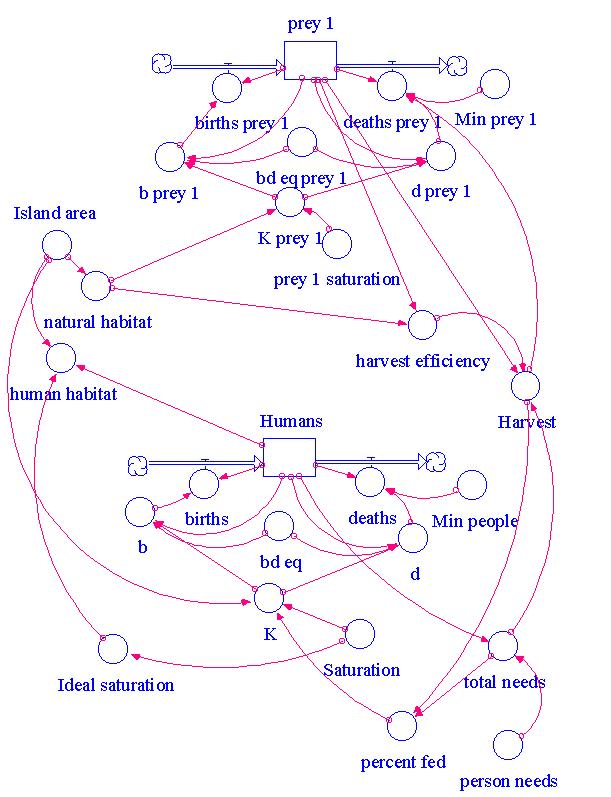
As in all mathematical modeling efforts, we begin with a grossly simplified set of relationships.† The first set of models consist of completely deterministic dynamic systems models.† We use the Stella simulation programming language to develop these simple models.
Simulation of an
island species.
Under the simplest circumstance, islands are not occupied by humans, so that straight-forward density-dependent population growth occurs.† The basic density-dependence model differs from the logistic equation to give us the flexibility of density and harvesting effects on either the birth rate, the death rate, or both.† Instead of specifying vital rates as a maximal growth rate (as does the logistic), our model specifies vital rates at demographic saturation (when DN = 0) because these values can be empirically determined for natural populations.
For a single mammalian island species (which we term prey 1), we define a saturation density,
![]() , which is the equilibrium density in individuals per
area.† The vital rates at saturation are
, which is the equilibrium density in individuals per
area.† The vital rates at saturation are
![]() †(birth rate) and
†(birth rate) and ![]() †(death rate) and
are constrained so that
†(death rate) and
are constrained so that ![]() =
=![]() .† The annual
birth rate at time t is given as
.† The annual
birth rate at time t is given as ![]() , where Nt
is the number of individuals at time t
and Ht is the habitable
island area (which may change with t
when humans begin utilizing some of the island area).† The annual death rate at time t
is
, where Nt
is the number of individuals at time t
and Ht is the habitable
island area (which may change with t
when humans begin utilizing some of the island area).† The annual death rate at time t
is ![]() .† The annual
change in population is Nt(btñdt) for Nt
greater than some minimum sustainable population size Nmin and -Nt
otherwise.
.† The annual
change in population is Nt(btñdt) for Nt
greater than some minimum sustainable population size Nmin and -Nt
otherwise.
The effect of humans on the ecosystem and the effect of the ecosystem on humans is a complex set of interactions.† We add humans to the basic model by assuming that they also have a saturation population density and vital rates at saturation.† The direct effects of humans on the ecosystem is to reduce the habitable area for the island prey species (but not the marine prey species), and they harvest a number of individuals each year, thus annually harvesting some number of individuals proportional to the human population size.
The human population size and growth is also affected by prey species.† We model dependence on island prey (particularly during the Jamon period) at low prey densities by an annual harvest requirement per person.† Thus, for some combinations of parameters, the prey will go extinct, followed by a crash in the human population size (or by falling below a threshold of minimum size required for sustaining a settlement).† A major difference between dependence on island prey and dependence on marine prey is that the human population does not reduce the habitable area for a marine species but does for an island species.
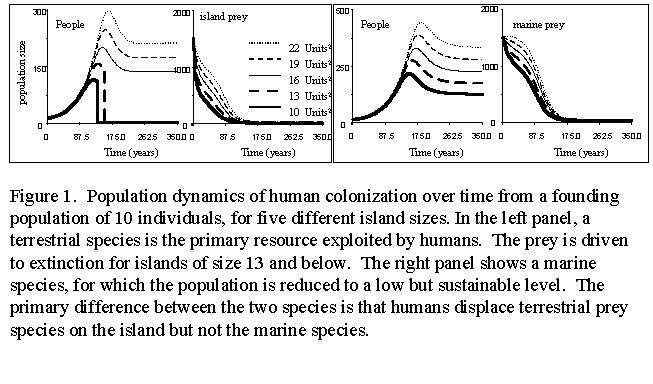
In Figure 1 , we plot population sizes for humans and a single island prey for a series of island ranging from a size of 6 to 15 (in arbitrary units for this uncalibrated model). (The Stella model is shown here ). † Under the parameterizations used here, a non-trivial stable equilibrium is reached for humans and prey for islands over 18, and prey are reduced to a level that is just capable of sustaining the human population.†† The effect of switching from terrestrial species to marine species can also be seen in the second set of curves.† During the later periods, there is a reduced dependence on terrestrial resources and marine resources.† We have explored a number of these models in the Stella programming language, which provides deterministic results for any given scenario.† Stochastic versions of the models will be used for estimation of parameters, as described in the statistical methods section below.† These changes in resource utilization and technology over time, and the concurrent effects on the ecosystem will provide the raw observations needed to estimate statistical models.
{kind=link}
{kind=link}
A discussion of some of the statistical methods can be found here .
Other stuff
∑† References
∑† links