|
Our visual systems perform all kinds of amazing jobs, from finding
constellations in the night sky, to picking out just the right strawberry
in the supermarket, to tracking a fly ball into a waiting glove. How do
our eyes and brains recognize shape, movement, depth, and color? How do
we so easily pick a friend's face out of a crowd, yet get fooled by
optical illusions? In this first of three units on the Sense of Sight, we
consider the anatomy and physiology of the eye, especially the retina, and
the initial pathways visual information takes to the brain. Part 2
discusses how various aspects of a visual scene are processed at higher
levels, and Part 3 delves into color vision.
1. Our eyes allow us to perceive
electromagnetic radiation reflected from objects
Most animals and many plants are photosensitive; that is, they can detect
different light intensities. Some organisms accomplish this with single
cells or with simple eyes that do not form images but do allow the
organism to react to light by moving toward or away from it. In order for
an eye to transmit more information about the world, however, it must have
a way of forming an image, a representation of the scene being viewed.
Higher invertebrates and virtually all vertebrates have complex,
image-forming eyes, and we will "focus" on the refracting eye found in the
octopus and in all vertebrates. Arthropods have compound eyes, which have
greater depth of focus than refracting eyes, but which sacrifice resolving
power or acuity. Our eyes, like those of many animals, detect a just
narrow range of all the wavelengths of electromagnetic radiation, that
between 380 and 760 nanometers. This range of light is called the visible
spectrum. Figure 1 shows how the visible spectrum fits into the entire
electromagnetic spectrum.
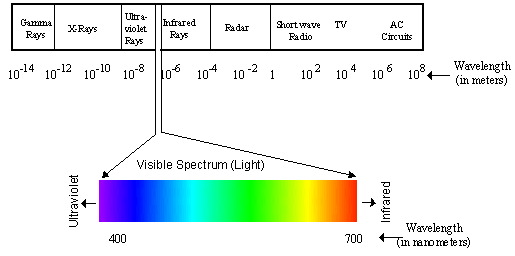
Figure 1. The electromagnetic spectrum and the visible spectrum.
2. The eyeball is an
optical device for focusing light
The mammalian eyeball (Figure 2) is an organ that focuses a visual scene
onto a sheet of specialized neural tissue, the retina, which lines the
back of the eye. Light from a scene passes through the cornea, pupil, and
lens on its way to the retina. The cornea and lens focus light from
objects onto photoreceptors, which absorb and then convert it into
electrical signals that carry information to the brain. Two pockets of
transparent fluid nourish eye tissues and maintain constant eye shape:
these are the aqueous and vitreous humors, through which the light also
passes. The lens projects an inverted image onto the retina in the same
way a camera lens projects an inverted image onto film; the brain adjusts
this inversion so we see the world in its correct orientation. To control
the images that fall upon our retinas, we can either turn our heads or
turn our eyes independently of our heads by contracting the extraocular
muscles, six bands of muscles that attach to the tough outside covering,
or sclera, of the eyeball and are innervated by cranial nerves. See Table
1 for a brief list of eyeball components and their functions.
The cornea and lens bend or refract light rays as they enter the eye, in
order to focus images on the retina. The eye can change the extent to
which rays are bent and thus can focus images of objects that are various
distances from the observer, by varying the curvature of the lens. The
ciliary muscle accomplishes this by contracting to lessen tension on the
lens and allowing it to round up so it can bend light rays more, or
relaxing for the opposite effect. This ciliary muscle is smooth or
non-voluntary muscle-you cannot "decide" to contract or relax it as you do
the skeletal muscle in a finger or facial muscle.
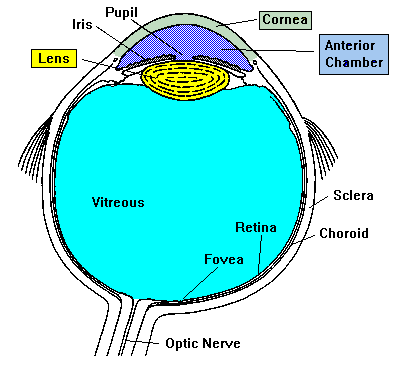
Figure 2. The mammalian eyeball.
3. Refractive errors in
the eye cause focusing problems
Refractive errors occur when the bending of light rays by the cornea and
lens does not focus the image correctly onto the retina. An eyeball that
is too long or too short for the optics of the cornea and lens or an
irregularly shaped cornea can cause refractive errors, which include
myopia (near-sightedness), hyperopia (far-sightedness), and astigmatism.
Myopia results either when the eyeball is too long or when the cornea is
curved too much, and the focused image falls in front of the retina.
Hyperopia is the opposite, with the image falling behind the retina.
Astigmatism results from a cornea that is not spherical. Fortunately,
most refractive errors can be corrected with prescription lenses.
| TABLE
1. PARTS OF THE EYE |
| STRUCTURE | FUNCTION |
| Aqueous humor | clear watery fluid found in the anterior
chamber of the eye; maintains pressure and nourishes the cornea and
lens |
| Vitreous humor | clear, jelly-like fluid found in the back
portion of the eye: maintains shape of the eye and attaches to the
retina |
| Blind spot | small area of the retina where the optic nerve
leaves the eye: any image falling here will not be seen |
| Ciliary muscles | involuntary muscles that change
the lens shape to allow focusing images of objects at different
distances |
| Cornea | transparent tissue covering the front of the eye:
does not have blood vessels; does have nerves |
| Cones | photoreceptors responsive to color and in bright
conditions; used for fine detail |
| Rods | photoreceptors responsive in low light conditions;
not useful for fine detail |
| Fovea | central part of the macula that provides sharpest
vision; contains only cones |
| Iris | circular band of muscles that controls the size of
the pupil. The pigmentation of the iris gives "color" to the eye. Blue
eyes have the least amount of pigment; brown eyes have the most |
| Lens | transparent tissue that bends light passing through
the eye: to focus light, the lens can change shape |
| Macula | small central area of the retina that provides
vision for fine work and reading |
| Optic nerve | bundle of over one million axons from ganglion
cells that carry visual signals from the eye to the brain |
| Pupil | hole in the center of the eye where light passes
through |
| Choroid | Thin tissue layer containing blood vessels,
sandwiched between the sclera and retina; also,
because of the high melanocytes content, the choroid acts as a
light-absorbing layer. |
| Retina | layer of tissue on the back portion of the eye
that contains cells responsive to light (photoreceptors) |
| Sclera | tough, white outer covering of the eyeball;
extraocular muscles attach here to move the eye |
4. The retina originates
from the brain and contains photoreceptors for
detecting light
The eye is formed during embryonic development by a combination of head
ectoderm and neural tube tissue, the latter forming the retina. Thus, the
retina is not a peripheral sensory organ like skin touch receptors or
taste buds on the tongue, but rather it is an outgrowth of central nervous
tisse. Because of this origin, the retina has layers of neurons, internal
circuits, and transmitters characteristic of the brain: it is a bit of
the brain that has journeyed out, literally, to have a look at the
environment.
The photoreceptors in the retina are of two types: rods and cones, so
named because of their shapes. These cells are actually specialized
neurons that detect light. Embedded in stacks of cell membranes in the
distal portions of rods and cones are molecules that absorb certain
wavelengths of light. These molecules are called photopigments and are
composed of two parts: a large trans-membrane protein, an opsin, and a
smaller chromophore, which is a metabolite of Vitamin A called
11-cis-retinal. The chromophore, which is embedded in the opsin, absorbs
light; in so doing it undergoes a shape change. This shape change in
turn activates the opsin, setting off a cascade of events that leads to a
change in the electrical state of a rod or cone cell membrane. This
change in the rod or cone cell membrane is then conducted via the rod or
cone axon to other neurons in the retina, and from there to the brain.
5. Rods function in dim light
In dim light, we use our rods, which cannot work in bright light. Rods
outnumber cones (120 million rods and about 6 million cones in each
retina) and they amplify a light signal much more than cones. Scientists
have demonstrated that absorption of even a single quantum (or photon) of
light can trigger a chromophore shape change in one molecule of rhodopsin
in a rod, leading to signal transmission. For transmission to occur, this
initial tiny event must be amplified: the activated molecule of rhodopsin
converts several thousand molecules of the next enzyme in the cascade to
the active form, and this amplification continues until the electrical
potential of the cell membrane changes and neurotransmitter release is
affected. Cones, on the other hand, must each absorb hundreds of photons
in order to send signals.
Another retinal mechanism that helps us to see in dim light or to see a
tiny amount of light in the dark is the convergence of rod cell signals
onto other retinal neurons. Many rods (up to 150) synapse onto the same
target neurons, where the signals are pooled and reinforce one another,
increasing the ability of the brain to detect a small amount of light. (A
synapse is a contact between a neuron and another cell where an
electrochemical signal [most commonly] is transmitted to the second cell.)
This convergence amplifies weak signals, but spatial resolution is lost
because rod responses are averaged. That is, we cannot see fine
detail using rods.
In order for our eyes to make the transition to dim light, rods must adapt
after being saturated with light in brighter conditions. Dark adaptation
of rods takes seven to ten minutes: during this time rhodopsin molecules,
in which the chromophore components have changed to the activated state,
return to the non-activated state so that they are able once again to
register changes in illumination. Other changes also occur in adaptation
to dark or dim conditions, including enlarging or dilating of the pupil,
which is controlled by the autonomic nervous system.
6. Cones mediate day
vision
Our vision in bright or moderate light is completely mediated by cones,
which provide color vision, black and white vision, and high acuity, the
ability to discern fine detail. Like rods, cones contain an opsin and the
chromophore 11-cis-retinal, but the opsins differ from rhodopsin so that
each cone is responsive to one of three colors: red, green or blue.
Cones are spread throughout the retina but are especially concentrated in
a central area called the macula. At the center of the macula is the
fovea, where only cones (no rods) are found, and these are densely packed.
When we want to read or inspect fine detail, we move our heads and eyes
until the image of interest falls onto the fovea. Because the fovea lacks
rods, it is easier to see in dim light by looking to the side of an object
instead of directly at it. You can test this by looking to the side of a
faint star so that its image falls on rods, rather than on the fovea where
it probably will not register. When you look directly at the faint star,
it disappears. In contrast to the wiring of rods, only a few cones
converge onto other retinal neurons to average their signals, so cones
provide better spatial resolution. In fact, each cone in the fovea
synapses onto only one neuron in the next relay in the retina. This gives
this area the ability to transmit fine detail, such as we use in reading.
Thus, cones mediate day vision and rods take over in dim light and at
night. Both rods and cones can operate at the same time under some
conditions: in dim or dark conditions, rods are most sensitive, but cones
respond to stimuli that are sufficiently bright. This is why we can see
the colors of neon lights on dark nights.
7. Visual information
travels from retinal ganglion cells to the brain
After converting light into electrical signals in their cell membranes,
rods and cones transmit this information to other neurons in internal
circuits in the retina for processing. From these cells, messages go to
the final retinal station, the ganglion cells, whose axons exit the
eyeball at the optic disc and form the optic nerve, which contains about
one million axons. Because all the nerve fibers converge at the optic
disc, no rods or cones are in this area and it forms a "blind spot" on the
retina: this may be easily demonstrated in a
classroom activity.
Within the optic nerve, a defined group of axons from each eye crosses
over to join the opposite optic nerve at the optic chiasma (see Figure 3),
so each side
of the brain receives visual information from both eyes. After the
chiasma, retinal axons go to one of three areas: two of these are in the
midbrain and one is in the thalamus. The information going to the
midbrain does not reach conscious levels but rather produces pupillary
reflexes (which are controlled by the autonomic nervous system) and eye
movements. In the thalamus, ganglion cell axons transmit signals to
neurons in the lateral geniculate nucleus (LGN) where information is
processed and then carried by LGN axons to the primary visual cortex in
the occipital lobe of the cerebrum. These cortical cells then send
messages to other "higher" cortical areas. Figure 3 shows the anatomy of
this system (the midbrain areas are not shown here).
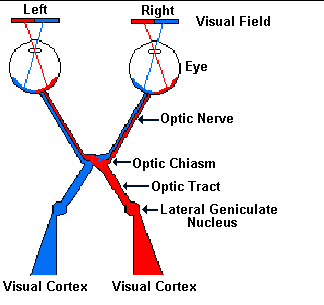
Figure 3. The visual pathway
8. We have an area of
central or focused vision and an area of peripheral
vision within our fields of vision
The visual field is defined as the view seen by the two eyes while looking
straight ahead (Figure 4). Without moving eyes or head, a person can see
details (well enough to read) within a limited angle drawn from a point
between the eyes on the forehead and two experimentally determined points
to the left and right in front of the viewer, at proper focal distance.
In addition to this area of clear or central vision, we can see objects
and movements to the sides of our heads, although as the distance around
to the sides increases, it becomes more difficult to identify objects.
The area of central vision includes objects whose images fall onto the
central area of the retina, the macula, and especially the fovea (defined
above). Cones in all other areas of the retina are in the periphery, and
while they convey visual information, they do not provide the resolving
power of the densely packed fovea.
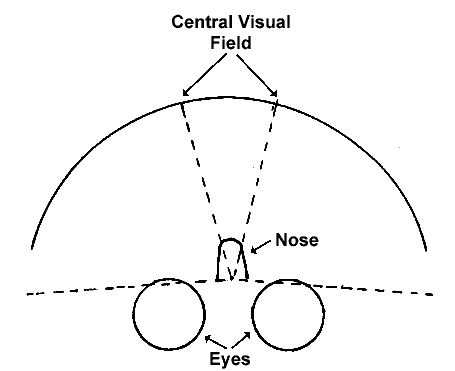
Figure 4. Complete visual field and central visual field, looking
down
onto the head. The complete visual field is the entire area in front of
the eyes from the end of one lateral dashed line to the other (including
the central visual field).
In addition to speaking of a central and a peripheral field of vision, we
can divide these fields by a vertical line down the middle into right and
left visual fields. Because of the way ganglion cell axons cross at the
optic chiasma, information from the entire right visual field (to the
right of a vertical line) goes to the left LGN, and from the left LGN, all
axons go to the left occipital cortex (Figure 3). Similarly, all left
visual field information goes to the right occipital cortex. Remember that
although each visual cortex receives information only from the opposite
visual field, this information is collected by defined parts of
both eyes.
9. Projections from the
retina to the brain generate retinotopic maps
As in the touch sensory system (and to some extent, other sensory
systems), visual information is mapped in an orderly fashion onto neurons
in the LGN of the thalamus. Further, this topographic mapping continues
when LGN neurons carry signals to the visual cortex. As in the touch
system, the mapping of the visual field is not isometric; that is, not
every area of the visual field is represented in proportion to its size.
Rather, the density of sensory neurons in a given area of the retina
determines how many central neurons are devoted to that retinal area, as
in the touch system where fingertips and lips have a much larger
representation in the parietal cortex than do trunk and arms. In the LGN
and primary visual cortex, about half of the neurons receive input from
the fovea (the eye's "fingertips") and area just around it, where cones
are densely packed and visual acuity is highest.
10. Defined groups of neurons in the primary
visual cortex process different aspects of visual
information
Several attributes of visual information go to the primary visual cortex:
motion, form or shape, and color. These aspects of the visual scene
travel to different modules or groups of cortical cells (some are given
names such as "columns" or "blobs.") In order for us to perceive and
interpret these kinds of visual information, other brain areas beyond the
primary visual cortex must process the signals and put the visual scene
back together.
11. Problems in different parts of the visual
system can cause blindness
People who lose cone vision are legally blind, whereas loss of only rod
function results in night blindness. Legal blindness is defined as 20/200
vision or worse; that is, a person is considered legally blind if he or
she must be
20 feet away to see an object that a person with normal vision can see at
200 feet. Some forms of blindness result from damage to both rods and
cones, while others originate with problems in different parts of the
visual system. For example, people with damage to particular parts of
the cerebral cortex lose specific aspects of vision, such as ability to
see parts of the visual field, or to perceive motion, or to recognize
faces. More information on these types of visual defects is given in Part
2 of this unit on the Sense of Sight.
|

 [Back to Top]
[Back to Top]![[email]](./gif/menue.gif)


