| The Role of Plant Competition During Early, Secondary Succession: An Experimental
Approach |
|
Photos
from the Starrbright study site where we are conducting long-term
species removal experiments to examine the role of interspecific competition
during the early stages of succession in the Pacific Northwest. |
 |
 |
||
| Broadcast burning of the study site, September 11, 1991 |
One
day after broadcast burning. Rebar posts mark the locations of experimental
plots. |
||
 |
 |
||
|
Ground
surface conditions in a 1 x 1 m experimental plot shortly after broadcast
burning. |
A "community
removal" plot 3 yr after burning. Note the numerous dead stems of Senecio sylvaticus (center) which dominated the previous year (see Abstract below). |
||
 |
 |
||
|
An
experimental block, 3 yr after burning. Dominant residual (resprouting) species is Rhododendron macrophyllum; dominant early seral species is the short-lived perennial, Gnaphalium microcephalum. |
Sampling
plant species density and cover, 7 yr after broadcast burning. Flowering
stems are Crepis capillaris. |
| From: Halpern,
C. B., J. A. Antos, M. A. Geyer, and A. M. Olson. 1997. Species replacement
during early secondary succession:
the abrupt decline of winter annual.
Ecology 78:621-631. (.pdf
file) |
|
|
|
|
|
Temporal changes in the performance of Senecio sylvaticus in species removal and control treatments for three growing seasons after broadcast burning (downward arrow). Population density, cover, and biomass peak in year 2, but removing some or all competitors does not affect the population decline in year 3. |
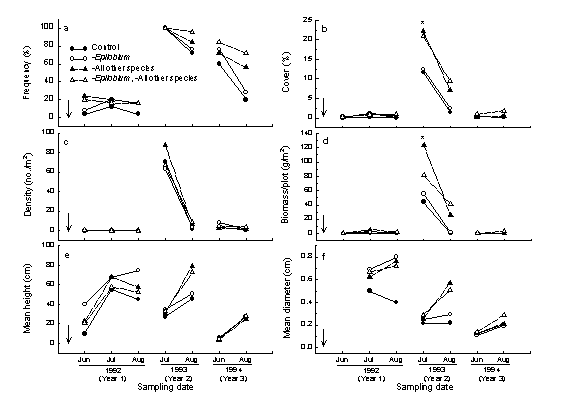
|
|
|
|
 |
 |
||
| Root system
of a 2-yr-old individual of Gnaphalium microcephalum, |
Root
system of a 2-yr-old individual of Epilobium angustifolium,
a long-lived perennial herb |
|
|
|
Before harvest, surface soils averaged total C (7.2 percent), total N (0.19 percent), extractable NH4+-N (5.2 ug/g), extractable NO3--N (0.19 ug/g), and pH (5.3). Samples collected 9 months after burning showed a 26 percent decline in concentration of total C. As organic matter burns, C is volatilized, lost as particulates or converted to inorganic carbon. However, addition of C to the surface soil also is possible if charcoal particles from burned duff or logging slash are leached into the soil. Because we used the dry-combustion analytical method that does not distinguish between organic and inorganic forms of C, the decline we observed is a minimum estimate of loss of organic C, which was likely greater, but partly offset by addition of charcoal. Concentrations of total N changed little after logging and burning, but extractable mineral N showed a fivefold increase 9 months after burning followed by an equally dramatic decline to preburn values in year 2. The coefficient of variation in extractable mineral N increased more than twofold after burning, which, together with the fivefold increase in the mean, yielded extremely high variation in available N after fire. In contrast to extractable mineral N, soil pH remained elevated through final sampling in 1994. Changes in concentration of extractable mineral N were weakly, but significantly, correlated with preharvest concentrations, but were unrelated to burn severity. Increases were observed in nearly all plots, and relatively large increases of extractable N were found in plots that had low, as well as high, initial concentrations. Nine months after burning (1992), concentration of extractable mineral N showed no relation to total plant biomass, which was invariably low during the first postburn growing season. Two and 3 years after burning, however, extractable mineral N showed a significant and increasingly strong negative relation with plant biomass (which had increased substantially), suggesting that N availability was measurably reduced by plant uptake. Most
variation in soil C and N before harvest occurred at small spatial
scales (within and among the 2 by 2 m plots); logging and broadcast
burning had little effect on this pattern. The large, but transient
increase, and the high spatial variation in available N after burning
may contribute to the rapid changes in abundance and the patchy spatial
distributions of early successional plants that benefit from catastrophic
disturbance. |
|
|
|
|
|
|
|
|